

Non-coding RNA Generated following Lariat Debranching Mediates Targeting of AID to DNA
- PMID: 25957684
- PMCID: PMC4426339
- DOI: 10.1016/j.cell.2015.03.020
Non-coding RNA Generated following Lariat Debranching Mediates Targeting of AID to DNA
Abstract
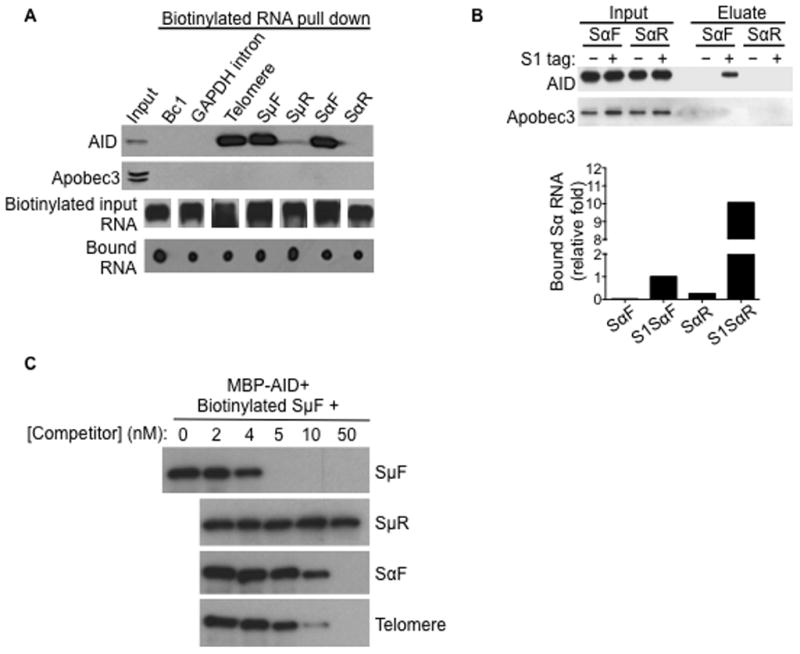
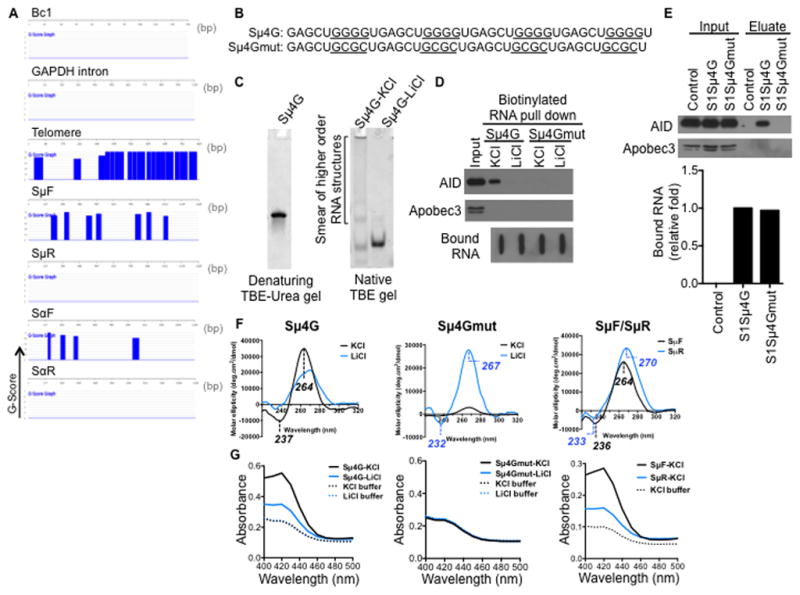
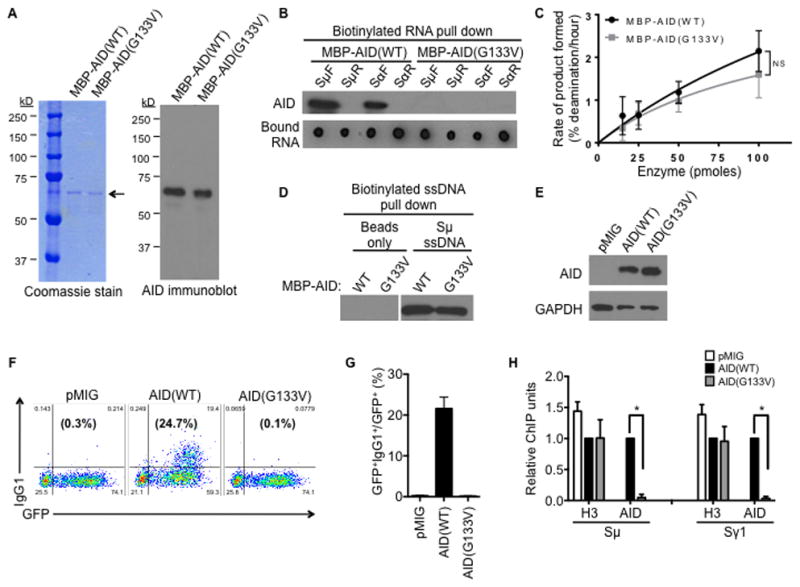
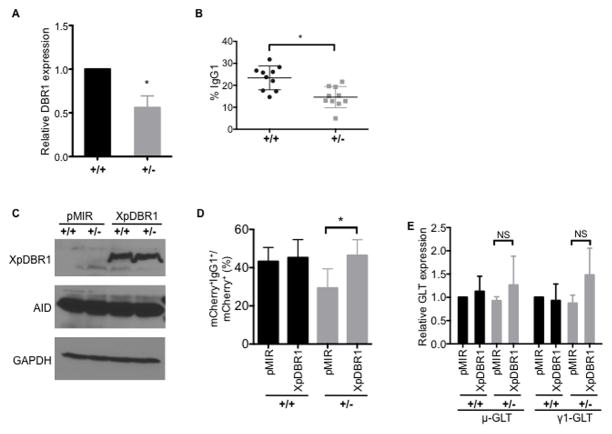
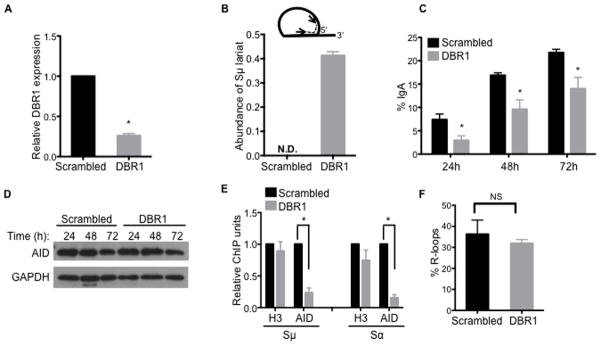
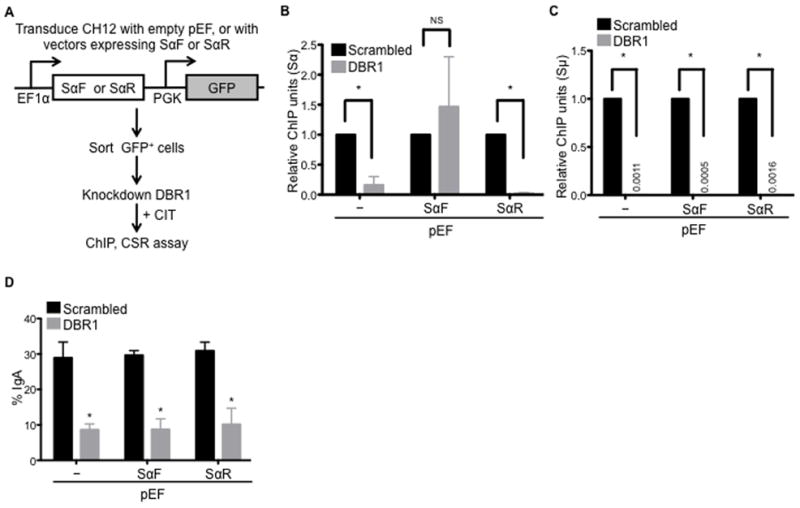
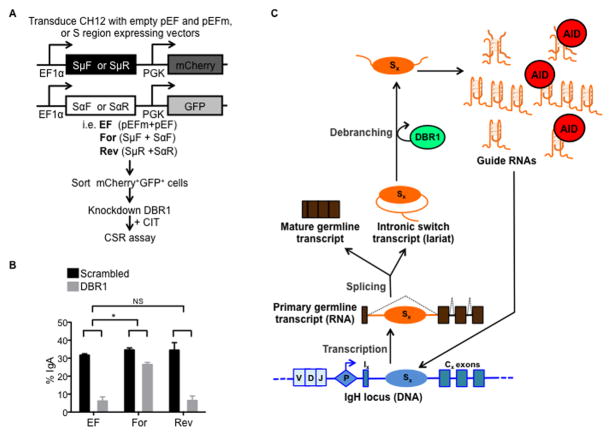
Transcription through immunoglobulin switch (S) regions is essential for class switch recombination (CSR), but no molecular function of the transcripts has been described. Likewise, recruitment of activation-induced cytidine deaminase (AID) to S regions is critical for CSR; however, the underlying mechanism has not been fully elucidated. Here, we demonstrate that intronic switch RNA acts in trans to target AID to S region DNA. AID binds directly to switch RNA through G-quadruplexes formed by the RNA molecules. Disruption of this interaction by mutation of a key residue in the putative RNA-binding domain of AID impairs recruitment of AID to S region DNA, thereby abolishing CSR. Additionally, inhibition of RNA lariat processing leads to loss of AID localization to S regions and compromises CSR; both defects can be rescued by exogenous expression of switch transcripts in a sequence-specific manner. These studies uncover an RNA-mediated mechanism of targeting AID to DNA.
Copyright © 2015 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare that they have no competing financial interests.
Figures
Similar articles
-
RNA Helicase DDX1 Converts RNA G-Quadruplex Structures into R-Loops to Promote IgH Class Switch Recombination.Mol Cell. 2018 May 17;70(4):650-662.e8. doi: 10.1016/j.molcel.2018.04.001. Epub 2018 May 3. Mol Cell. 2018. PMID: 29731414 Free PMC article.
-
Unfolding the Role of Splicing Factors and RNA Debranching in AID Mediated Antibody Diversification.Int Rev Immunol. 2021;40(4):289-306. doi: 10.1080/08830185.2020.1815725. Epub 2020 Sep 14. Int Rev Immunol. 2021. PMID: 32924658 Review.
-
AID- and Ung-dependent generation of staggered double-strand DNA breaks in immunoglobulin class switch DNA recombination: a post-cleavage role for AID.Mol Immunol. 2008 Nov;46(1):45-61. doi: 10.1016/j.molimm.2008.07.003. Epub 2008 Aug 28. Mol Immunol. 2008. PMID: 18760480 Free PMC article.
-
A Hyper-IgM Syndrome Mutation in Activation-Induced Cytidine Deaminase Disrupts G-Quadruplex Binding and Genome-wide Chromatin Localization.Immunity. 2020 Nov 17;53(5):952-970.e11. doi: 10.1016/j.immuni.2020.10.003. Epub 2020 Oct 23. Immunity. 2020. PMID: 33098766 Free PMC article.
-
Regulating infidelity: RNA-mediated recruitment of AID to DNA during class switch recombination.Eur J Immunol. 2016 Mar;46(3):523-30. doi: 10.1002/eji.201545809. Epub 2016 Feb 22. Eur J Immunol. 2016. PMID: 26799454 Free PMC article. Review.
Cited by
-
Regulation of long non-coding RNAs and genome dynamics by the RNA surveillance machinery.Nat Rev Mol Cell Biol. 2020 Mar;21(3):123-136. doi: 10.1038/s41580-019-0209-0. Epub 2020 Feb 4. Nat Rev Mol Cell Biol. 2020. PMID: 32020081 Free PMC article. Review.
-
AID in Antibody Diversification: There and Back Again.Trends Immunol. 2020 Jul;41(7):586-600. doi: 10.1016/j.it.2020.04.009. Epub 2020 Apr 27. Trends Immunol. 2020. PMID: 32434680 Free PMC article. Review.
-
The debranching enzyme Dbr1 regulates lariat turnover and intron splicing.Nat Commun. 2024 May 30;15(1):4617. doi: 10.1038/s41467-024-48696-1. Nat Commun. 2024. PMID: 38816363 Free PMC article.
-
Ago2 and a miRNA reduce Topoisomerase 1 for enhancing DNA cleavage in antibody diversification by activation-induced cytidine deaminase.Proc Natl Acad Sci U S A. 2023 May 2;120(18):e2216918120. doi: 10.1073/pnas.2216918120. Epub 2023 Apr 24. Proc Natl Acad Sci U S A. 2023. PMID: 37094168 Free PMC article.
-
R Loops in the Regulation of Antibody Gene Diversification.Genes (Basel). 2017 Jun 2;8(6):154. doi: 10.3390/genes8060154. Genes (Basel). 2017. PMID: 28574479 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
