

Comment
doi: 10.1016/j.molcel.2015.04.003.
Calcium signals tune the fidelity of transcriptional responses
Affiliations
- PMID: 25884365
- PMCID: PMC4743736
- DOI: 10.1016/j.molcel.2015.04.003
Item in Clipboard
Comment
Calcium signals tune the fidelity of transcriptional responses
Mol Cell.
.
Abstract
In this issue of Molecular Cell, Kar and Parekh (2015) reveal the remarkable intricacy and accuracy of Ca(2+) signals in differentially controlling the function of closely related transcription factors.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
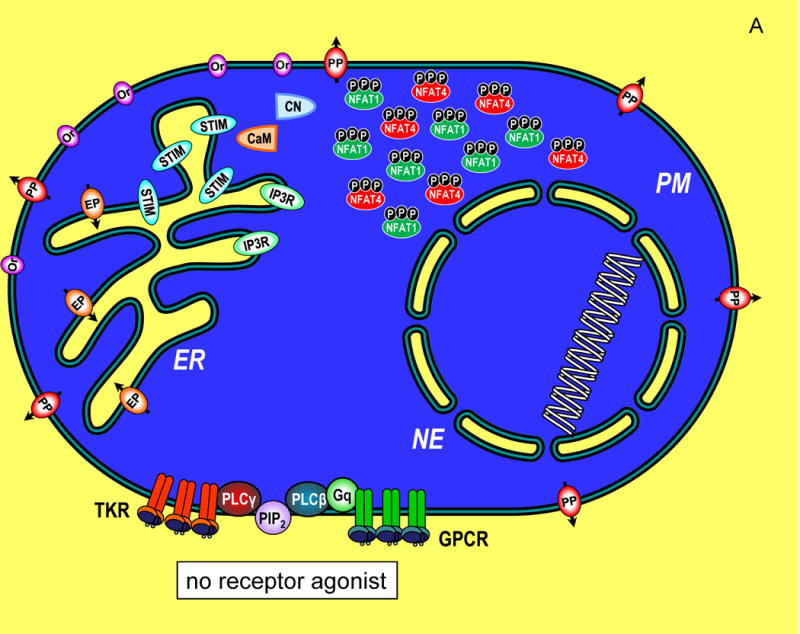
Transcriptional control through integrated Ca2+ signal
transduction pathways. (A) In a resting cell, cytoplasmic
Ca2+ is maintained at approximately 100 nM (blue) by
endoplasmic reticulum (ER) Ca2+ pumps (EP) and plasma
membrane (PM) Ca2+ pumps (PP). Ca2+ levels
outside the cell and within the ER lumen and nuclear envelope (NE) are much
higher, approximately 1 mM (yellow). Components of the Ca2+
signaling pathway include tyrosine kinase receptors (TKR), phospholipase
C-γ (PLC-γ), G protein-coupled receptors (GPCR), G protein q
(Gq), phospholipase C-β (PLC-β), phosphatidylinositol
4,5-bisphosphate (PIP2), inositol 1,4,5-trisphosphate
(IP3), InsP3 receptor (IP3R), STIM proteins
(STIM), Orai channels (Or), calmodulin (CaM), calcineurin (CN), nuclear factor
of activated T cells (NFAT1 and NFAT4) shown in their cytoplasmic,
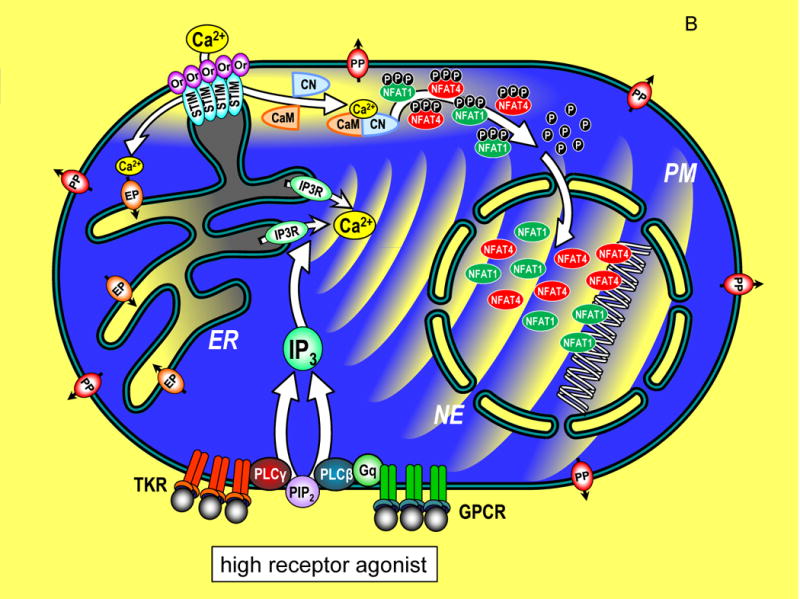
phosphorylated state. (B) A cell in which Ca2+ signals are
induced by maximal levels of TKR or GPCR agonists, inducing PIP2
breakdown to release high levels of InsP3 which activates large
pulsatile ER Ca2+ release through InsP3Rs,
manifested as global Ca2+ oscillations that traverse the
cytoplasm to activate cellular responses. The cytosolic Ca2+
pulses penetrate within the nucleoplasmic space through permeable nuclear
envelope pores. Ca2+ release from ER, depletes luminal stored
Ca2+ (grey shading). STIM proteins, widely distributed
across ER in the resting state, become activated and translocate into ER-PM
junctions where they contact the PM and expose an Orai channel binding domain
that tethers and gates Orai channels to allow Ca2+ to enter
the cytosol. The Orai-mediated Ca2+ entry provides highly
localized Ca2+ signals (within a few nm of the PM; yellow
shading). The entering Ca2+ binds to CaM and triggers CN
association and activates CN phosphatase activity which dephosphorylates NFAT1
and NFAT4 exposing nuclear import signal sequences. The dephosphorylated NFAT1
and NFAT4 molecules enter the nucleus wherein they each target distinct arrays
of genes. The continuation of Ca2+ signals induces retention
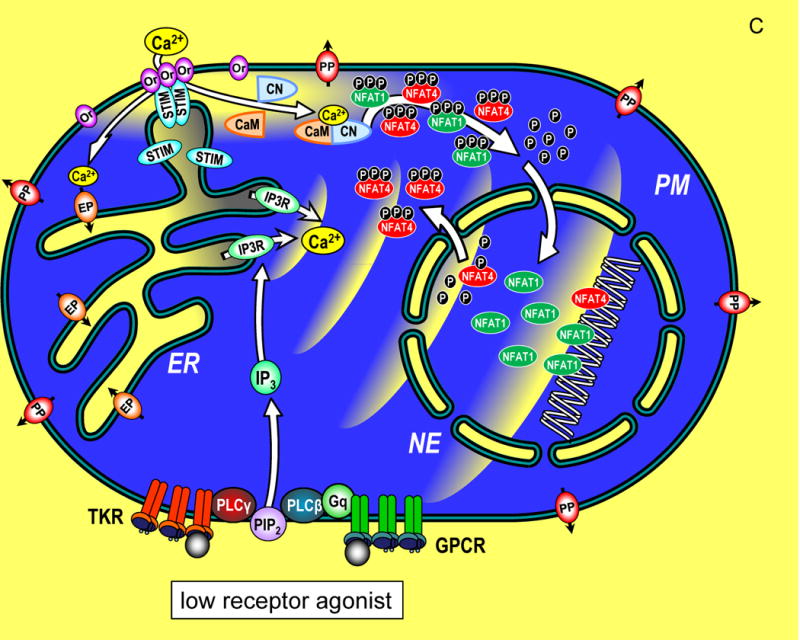
of NFAT molecules in the nucleus and prolonged gene expression. (C) A cell in
which submaximal levels of TKR or GPCR agonists induces modest InsP3
levels and smaller, less frequent pulses of Ca2+ in the
cytoplasm. STIM and Orai proteins are still activated, albeit to a lower extent,
and Ca2+ entry triggers NFAT1 and NFAT4 phosphorylation and
nuclear import. Within the nucleus, the smaller pulsatile rises in
Ca2+ are insufficient to retain NFAT4, but NFAT1 remains
activated in the nucleus despite the lower nucleoplasmic
Ca2+. The lower nuclear Ca2+ may
specifically trigger an increased rate of phosphorylation and faster export of
NFAT4 from the nucleus compared to NFAT1. Since NFAT1 and NFAT4 target different
genes, the pattern of transcription is tuned by both the Orai-mediated
Ca2+ entry, and the frequency of Ca2+
pulses released from ER.
Transcriptional control through integrated Ca2+ signal
transduction pathways. (A) In a resting cell, cytoplasmic
Ca2+ is maintained at approximately 100 nM (blue) by
endoplasmic reticulum (ER) Ca2+ pumps (EP) and plasma
membrane (PM) Ca2+ pumps (PP). Ca2+ levels
outside the cell and within the ER lumen and nuclear envelope (NE) are much
higher, approximately 1 mM (yellow). Components of the Ca2+
signaling pathway include tyrosine kinase receptors (TKR), phospholipase
C-γ (PLC-γ), G protein-coupled receptors (GPCR), G protein q
(Gq), phospholipase C-β (PLC-β), phosphatidylinositol
4,5-bisphosphate (PIP2), inositol 1,4,5-trisphosphate
(IP3), InsP3 receptor (IP3R), STIM proteins
(STIM), Orai channels (Or), calmodulin (CaM), calcineurin (CN), nuclear factor
of activated T cells (NFAT1 and NFAT4) shown in their cytoplasmic,
phosphorylated state. (B) A cell in which Ca2+ signals are
induced by maximal levels of TKR or GPCR agonists, inducing PIP2
breakdown to release high levels of InsP3 which activates large
pulsatile ER Ca2+ release through InsP3Rs,
manifested as global Ca2+ oscillations that traverse the
cytoplasm to activate cellular responses. The cytosolic Ca2+
pulses penetrate within the nucleoplasmic space through permeable nuclear
envelope pores. Ca2+ release from ER, depletes luminal stored
Ca2+ (grey shading). STIM proteins, widely distributed
across ER in the resting state, become activated and translocate into ER-PM
junctions where they contact the PM and expose an Orai channel binding domain
that tethers and gates Orai channels to allow Ca2+ to enter
the cytosol. The Orai-mediated Ca2+ entry provides highly
localized Ca2+ signals (within a few nm of the PM; yellow
shading). The entering Ca2+ binds to CaM and triggers CN
association and activates CN phosphatase activity which dephosphorylates NFAT1
and NFAT4 exposing nuclear import signal sequences. The dephosphorylated NFAT1
and NFAT4 molecules enter the nucleus wherein they each target distinct arrays
of genes. The continuation of Ca2+ signals induces retention
of NFAT molecules in the nucleus and prolonged gene expression. (C) A cell in
which submaximal levels of TKR or GPCR agonists induces modest InsP3
levels and smaller, less frequent pulses of Ca2+ in the
cytoplasm. STIM and Orai proteins are still activated, albeit to a lower extent,
and Ca2+ entry triggers NFAT1 and NFAT4 phosphorylation and
nuclear import. Within the nucleus, the smaller pulsatile rises in
Ca2+ are insufficient to retain NFAT4, but NFAT1 remains
activated in the nucleus despite the lower nucleoplasmic
Ca2+. The lower nuclear Ca2+ may
specifically trigger an increased rate of phosphorylation and faster export of
NFAT4 from the nucleus compared to NFAT1. Since NFAT1 and NFAT4 target different
genes, the pattern of transcription is tuned by both the Orai-mediated
Ca2+ entry, and the frequency of Ca2+
pulses released from ER.
Transcriptional control through integrated Ca2+ signal
transduction pathways. (A) In a resting cell, cytoplasmic
Ca2+ is maintained at approximately 100 nM (blue) by
endoplasmic reticulum (ER) Ca2+ pumps (EP) and plasma
membrane (PM) Ca2+ pumps (PP). Ca2+ levels
outside the cell and within the ER lumen and nuclear envelope (NE) are much
higher, approximately 1 mM (yellow). Components of the Ca2+
signaling pathway include tyrosine kinase receptors (TKR), phospholipase
C-γ (PLC-γ), G protein-coupled receptors (GPCR), G protein q
(Gq), phospholipase C-β (PLC-β), phosphatidylinositol
4,5-bisphosphate (PIP2), inositol 1,4,5-trisphosphate
(IP3), InsP3 receptor (IP3R), STIM proteins
(STIM), Orai channels (Or), calmodulin (CaM), calcineurin (CN), nuclear factor
of activated T cells (NFAT1 and NFAT4) shown in their cytoplasmic,
phosphorylated state. (B) A cell in which Ca2+ signals are
induced by maximal levels of TKR or GPCR agonists, inducing PIP2
breakdown to release high levels of InsP3 which activates large
pulsatile ER Ca2+ release through InsP3Rs,
manifested as global Ca2+ oscillations that traverse the
cytoplasm to activate cellular responses. The cytosolic Ca2+
pulses penetrate within the nucleoplasmic space through permeable nuclear
envelope pores. Ca2+ release from ER, depletes luminal stored
Ca2+ (grey shading). STIM proteins, widely distributed
across ER in the resting state, become activated and translocate into ER-PM
junctions where they contact the PM and expose an Orai channel binding domain
that tethers and gates Orai channels to allow Ca2+ to enter
the cytosol. The Orai-mediated Ca2+ entry provides highly
localized Ca2+ signals (within a few nm of the PM; yellow
shading). The entering Ca2+ binds to CaM and triggers CN
association and activates CN phosphatase activity which dephosphorylates NFAT1
and NFAT4 exposing nuclear import signal sequences. The dephosphorylated NFAT1
and NFAT4 molecules enter the nucleus wherein they each target distinct arrays
of genes. The continuation of Ca2+ signals induces retention
of NFAT molecules in the nucleus and prolonged gene expression. (C) A cell in
which submaximal levels of TKR or GPCR agonists induces modest InsP3
levels and smaller, less frequent pulses of Ca2+ in the
cytoplasm. STIM and Orai proteins are still activated, albeit to a lower extent,
and Ca2+ entry triggers NFAT1 and NFAT4 phosphorylation and
nuclear import. Within the nucleus, the smaller pulsatile rises in
Ca2+ are insufficient to retain NFAT4, but NFAT1 remains
activated in the nucleus despite the lower nucleoplasmic
Ca2+. The lower nuclear Ca2+ may
specifically trigger an increased rate of phosphorylation and faster export of
NFAT4 from the nucleus compared to NFAT1. Since NFAT1 and NFAT4 target different
genes, the pattern of transcription is tuned by both the Orai-mediated
Ca2+ entry, and the frequency of Ca2+
pulses released from ER.
Comment on
-
Distinct spatial Ca2+ signatures selectively activate different NFAT transcription factor isoforms.Mol Cell. 2015 Apr 16;58(2):232-43. doi: 10.1016/j.molcel.2015.02.027. Epub 2015 Mar 26. Mol Cell. 2015. PMID: 25818645 Free PMC article.
Similar articles
-
Store-operated calcium channels and pro-inflammatory signals.Acta Pharmacol Sin. 2006 Jul;27(7):813-20. doi: 10.1111/j.1745-7254.2006.00395.x. Acta Pharmacol Sin. 2006. PMID: 16787563 Review.
-
Signalling to transcription: store-operated Ca2+ entry and NFAT activation in lymphocytes.Cell Calcium. 2007 Aug;42(2):145-56. doi: 10.1016/j.ceca.2007.03.007. Epub 2007 Jun 18. Cell Calcium. 2007. PMID: 17572487 Review.
-
Calcium signaling mechanisms in T lymphocytes.Annu Rev Immunol. 2001;19:497-521. doi: 10.1146/annurev.immunol.19.1.497. Annu Rev Immunol. 2001. PMID: 11244045 Review.
-
The Ca(v)1.4 calcium channel is a critical regulator of T cell receptor signaling and naive T cell homeostasis.Immunity. 2011 Sep 23;35(3):349-60. doi: 10.1016/j.immuni.2011.07.011. Epub 2011 Aug 11. Immunity. 2011. PMID: 21835646
-
Regulation of phospholipase C-gamma2 networks in B lymphocytes.Adv Immunol. 2005;88:73-96. doi: 10.1016/S0065-2776(05)88003-4. Adv Immunol. 2005. PMID: 16227088 Review.
Cited by
-
The STIM1-binding site nexus remotely controls Orai1 channel gating.Nat Commun. 2016 Dec 8;7:13725. doi: 10.1038/ncomms13725. Nat Commun. 2016. PMID: 27929067 Free PMC article.
-
The STIM-Orai coupling interface and gating of the Orai1 channel.Cell Calcium. 2017 May;63:8-13. doi: 10.1016/j.ceca.2017.01.001. Epub 2017 Jan 8. Cell Calcium. 2017. PMID: 28087079 Free PMC article. Review.
-
Ca2+ Microdomains, Calcineurin and the Regulation of Gene Transcription.Cells. 2021 Apr 12;10(4):875. doi: 10.3390/cells10040875. Cells. 2021. PMID: 33921430 Free PMC article. Review.
-
Pore properties of Orai1 calcium channel dimers and their activation by the STIM1 ER calcium sensor.J Biol Chem. 2018 Aug 17;293(33):12962-12974. doi: 10.1074/jbc.RA118.003424. Epub 2018 Jun 28. J Biol Chem. 2018. PMID: 29954946 Free PMC article.
-
The STIM-Orai Pathway: Conformational Coupling Between STIM and Orai in the Activation of Store-Operated Ca2+ Entry.Adv Exp Med Biol. 2017;993:83-98. doi: 10.1007/978-3-319-57732-6_5. Adv Exp Med Biol. 2017. PMID: 28900910 Free PMC article. Review.
References
-
- Bading H. Nuclear calcium signalling in the regulation of brain function. Nature Rev Neuroscience. 2013;14:593–608. - PubMed
-
- Baine I, Abe BT, Macian F. Regulation of T-cell tolerance by calcium/NFAT signaling. Immunol Rev. 2009;231:225–240. - PubMed
-
- Berridge MJ, Bootman MD, Roderick HL. Calcium signalling: dynamics, homeostasis and remodelling. Nat Rev Mol Cell Biol. 2003;4:517–529. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
