

Microtubule minus end motors kinesin-14 and dynein drive nuclear congression in parallel pathways
- PMID: 25869666
- PMCID: PMC4395489
- DOI: 10.1083/jcb.201409087
Microtubule minus end motors kinesin-14 and dynein drive nuclear congression in parallel pathways
Abstract
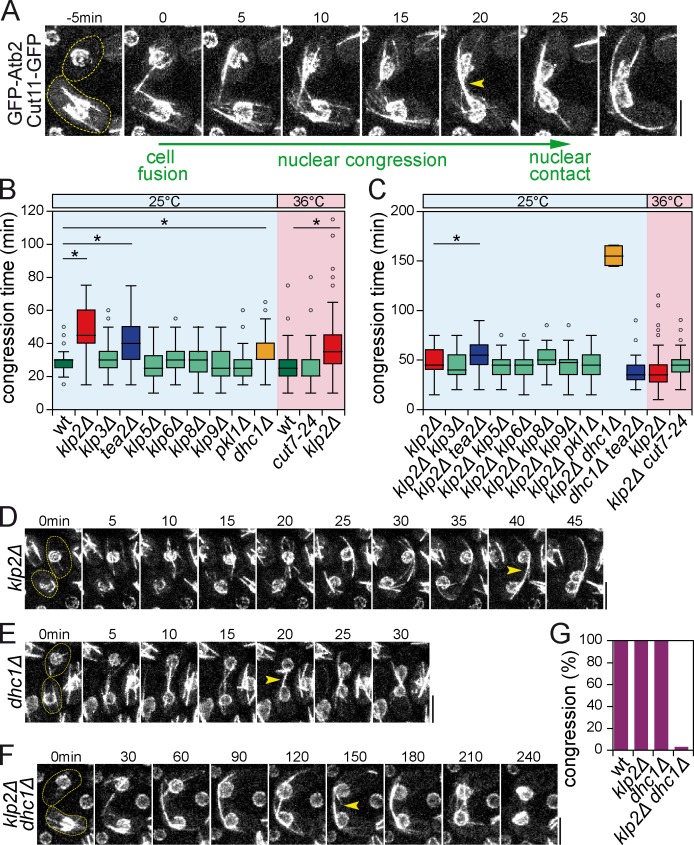
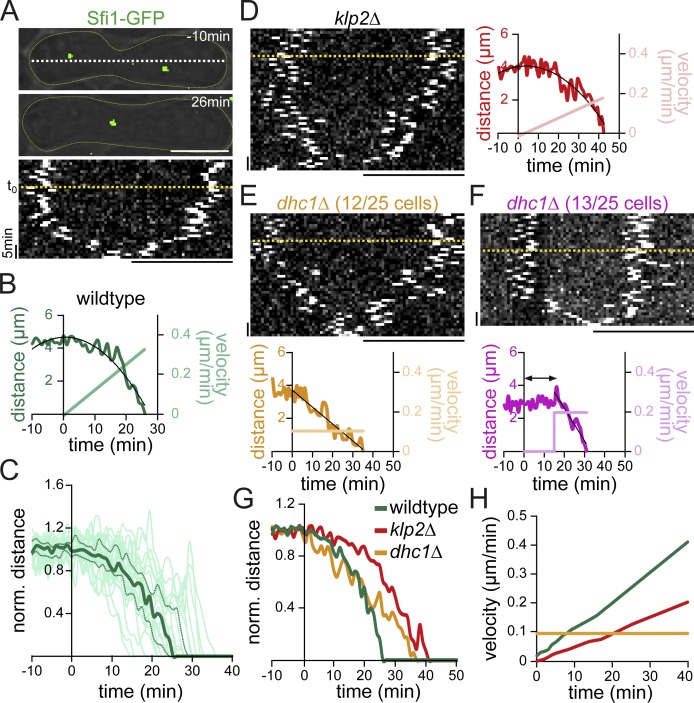
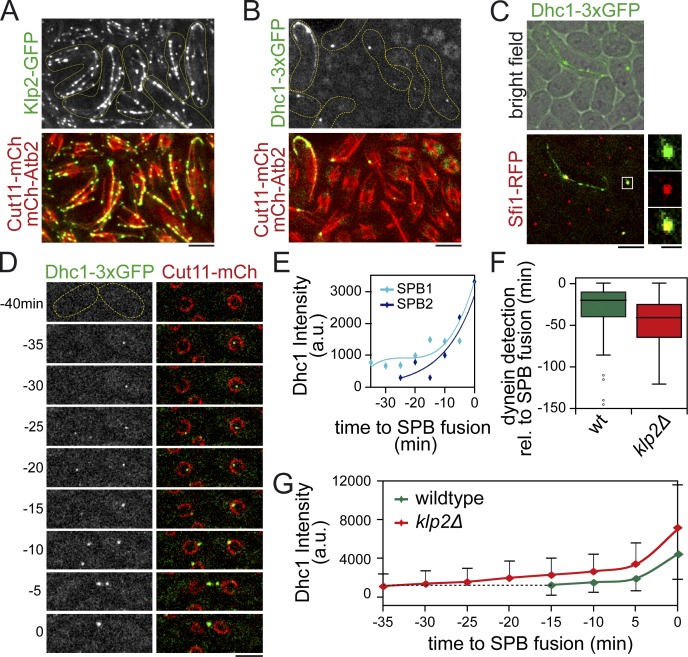
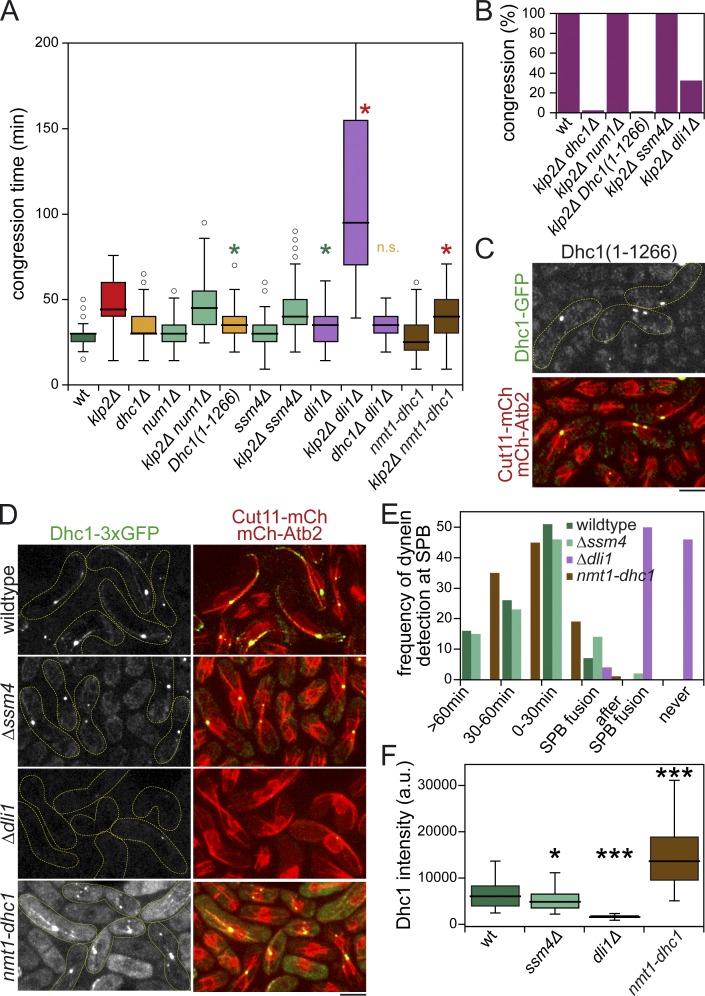
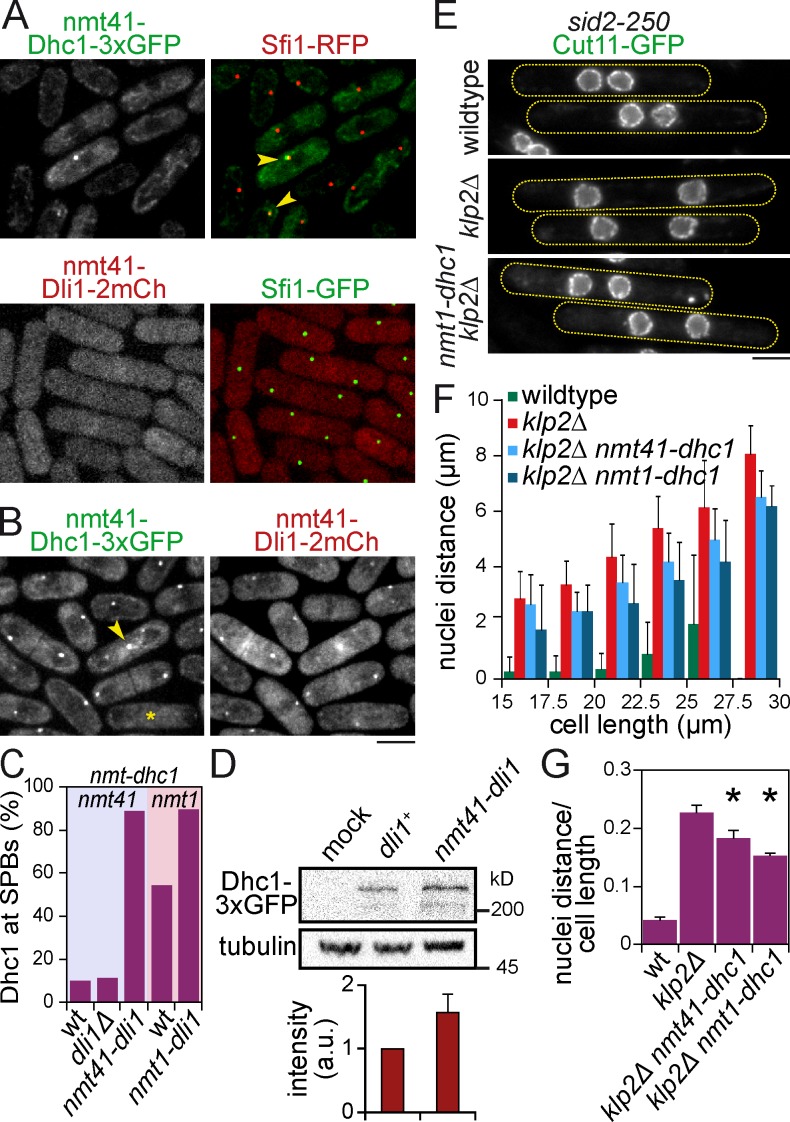
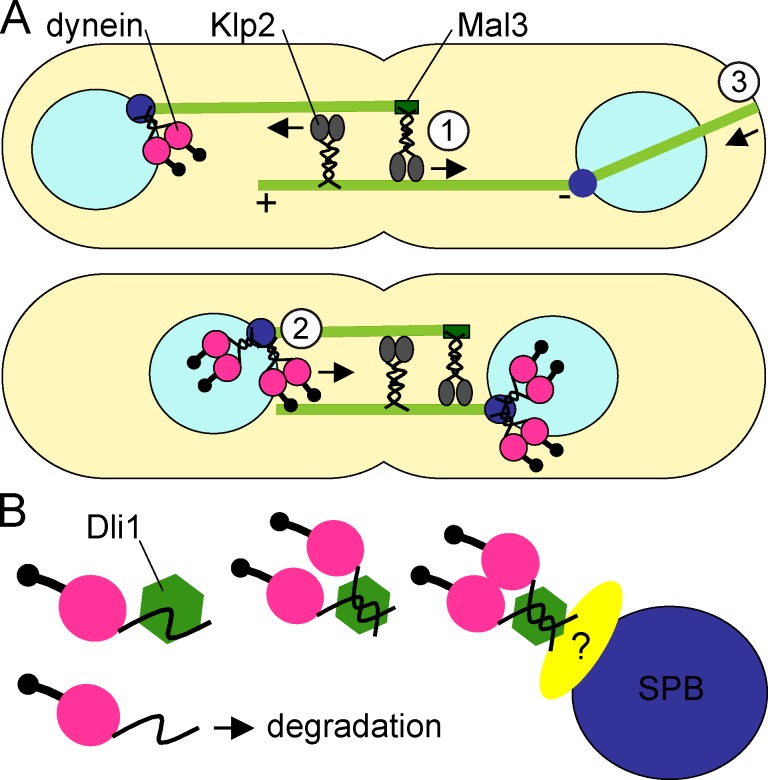
Microtubules (MTs) and associated motors play a central role in nuclear migration, which is crucial for diverse biological functions including cell division, polarity, and sexual reproduction. In this paper, we report a dual mechanism underlying nuclear congression during fission yeast karyogamy upon mating of haploid cells. Using microfluidic chambers for long-term imaging, we captured the precise timing of nuclear congression and identified two minus end-directed motors operating in parallel in this process. Kinesin-14 Klp2 associated with MTs may cross-link and slide antiparallel MTs emanating from the two nuclei, whereas dynein accumulating at spindle pole bodies (SPBs) may pull MTs nucleated from the opposite SPB. Klp2-dependent nuclear congression proceeds at constant speed, whereas dynein accumulation results in an increase of nuclear velocity over time. Surprisingly, the light intermediate chain Dli1, but not dynactin, is required for this previously unknown function of dynein. We conclude that efficient nuclear congression depends on the cooperation of two minus end-directed motors.
© 2015 Scheffler et al.
Figures
Similar articles
-
Mitotic chromosome biorientation in fission yeast is enhanced by dynein and a minus-end-directed, kinesin-like protein.Mol Biol Cell. 2007 Jun;18(6):2216-25. doi: 10.1091/mbc.e06-11-0987. Epub 2007 Apr 4. Mol Biol Cell. 2007. PMID: 17409356 Free PMC article.
-
Force by minus-end motors Dhc1 and Klp2 collapses the S. pombe spindle after laser ablation.Biophys J. 2022 Jan 18;121(2):263-276. doi: 10.1016/j.bpj.2021.12.019. Epub 2021 Dec 21. Biophys J. 2022. PMID: 34951983 Free PMC article.
-
Contribution of dynein light intermediate and intermediate chains to subcellular localization of the dynein-dynactin motor complex in Schizosaccharomyces pombe.Genes Cells. 2010 Apr 1;15(4):359-72. doi: 10.1111/j.1365-2443.2010.01386.x. Epub 2010 Mar 14. Genes Cells. 2010. PMID: 20298435
-
LIS1 at the microtubule plus end and its role in dynein-mediated nuclear migration.J Cell Biol. 2003 Feb 3;160(3):289-90. doi: 10.1083/jcb.200212168. J Cell Biol. 2003. PMID: 12566423 Free PMC article. Review.
-
Bidirectional transport of organelles: unity and struggle of opposing motors.Cell Biol Int. 2012 Jan;36(1):1-6. doi: 10.1042/CBI20110413. Cell Biol Int. 2012. PMID: 22142363 Review.
Cited by
-
A lumenal interrupted helix in human sperm tail microtubules.Sci Rep. 2018 Feb 9;8(1):2727. doi: 10.1038/s41598-018-21165-8. Sci Rep. 2018. PMID: 29426884 Free PMC article.
-
Automated tracking of S. pombe spindle elongation dynamics.J Microsc. 2021 Oct;284(1):83-94. doi: 10.1111/jmi.13044. Epub 2021 Jul 8. J Microsc. 2021. PMID: 34152622 Free PMC article.
-
Position matters: multiple functions of LINC-dependent chromosome positioning during meiosis.Curr Genet. 2017 Dec;63(6):1037-1052. doi: 10.1007/s00294-017-0699-2. Epub 2017 May 10. Curr Genet. 2017. PMID: 28493118
-
Nuclear movement in fungi.Semin Cell Dev Biol. 2018 Oct;82:3-16. doi: 10.1016/j.semcdb.2017.10.024. Epub 2017 Dec 11. Semin Cell Dev Biol. 2018. PMID: 29241689 Free PMC article. Review.
-
The mitotic kinesin-14 KlpA contains a context-dependent directionality switch.Nat Commun. 2017 Jan 4;8:13999. doi: 10.1038/ncomms13999. Nat Commun. 2017. PMID: 28051135 Free PMC article.
References
-
- Almonacid M., Celton-Morizur S., Jakubowski J.L., Dingli F., Loew D., Mayeux A., Chen J.S., Gould K.L., Clifford D.M., and Paoletti A.. 2011. Temporal control of contractile ring assembly by Plo1 regulation of myosin II recruitment by Mid1/anillin. Curr. Biol. 21:473–479 10.1016/j.cub.2011.02.003 - DOI - PMC - PubMed
-
- Bähler J., Wu J.Q., Longtine M.S., Shah N.G., McKenzie A. III, Steever A.B., Wach A., Philippsen P., and Pringle J.R.. 1998. Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast. 14:943–951 10.1002/(SICI)1097-0061(199807)14:10<943::AID-YEA292>3.0.CO;2-Y - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
