

Review
doi: 10.1186/s12915-015-0129-1.
Mitophagy and the mitochondrial unfolded protein response in neurodegeneration and bacterial infection
- PMID: 25857750
- PMCID: PMC4384303
- DOI: 10.1186/s12915-015-0129-1
Item in Clipboard
Review
Mitophagy and the mitochondrial unfolded protein response in neurodegeneration and bacterial infection
BMC Biol.
.
Abstract
Mitochondria are highly dynamic and structurally complex organelles that provide multiple essential metabolic functions. Mitochondrial dysfunction is associated with neurodegenerative conditions such as Parkinson's disease, as well as bacterial infection. Here, we explore the roles of mitochondrial autophagy (mitophagy) and the mitochondrial unfolded protein response (UPR(mt)) in the response to mitochondrial dysfunction, focusing in particular on recent evidence on the role of mitochondrial import efficiency in the regulation of these stress pathways and how they may interact to protect the mitochondrial pool while initiating an innate immune response to protect against bacterial pathogens.
Figures
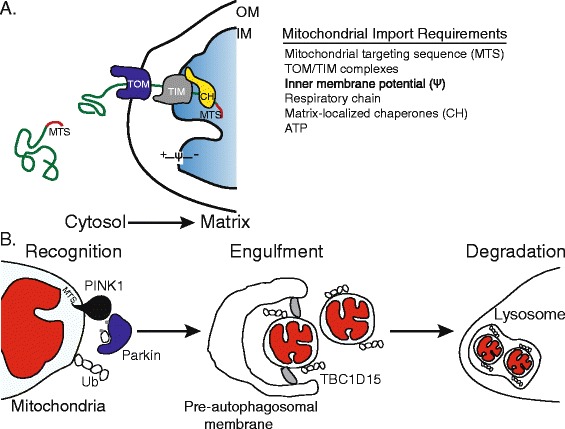
Mitochondrial protein import and PINK1-mediated mitophagy. (A) The vast majority of mitochondrial proteins are encoded by nuclear genes, synthesized on cytosolic ribosomes and targeted to mitochondria via mitochondrial targeting sequences (MTS). To reach the mitochondrial matrix, proteins synthesized on cytosolic ribosomes first interact with the translocase of the outer membrane (TOM) and then with the translocase of the inner membrane (TIM). Crossing the inner membrane requires both complexes, a membrane potential (Ψ) across the inner mitochondrial membrane that is generated by the respiratory chain, ATP and molecular chaperones (CH) within the mitochondrial matrix. Once in the matrix, the MTS is typically cleaved, allowing the protein to fold and assemble appropriately. Perturbations to the TOM/TIM complexes, respiratory chain, membrane potential and mitochondrial chaperones results in reduced mitochondrial import efficiency. (B) The kinase PINK1 serves to monitor mitochondrial health and initiate mitochondrial degradation when an organelle is severely damaged. Normally, PINK1, localized to mitochondria by its MTS sequence, is efficiently imported into the mitochondrion and subsequently degraded. However, when a mitochondrion is damaged (red), resulting in a depleted inner membrane potential or because of high levels of unfolded proteins in the matrix, PINK1 fails to be imported and accumulates on the mitochondrial outer membrane, allowing recognition of the damaged organelle in a sequence of steps, the first of which is the recruitment of the ubiquitin ligase Parkin to the outer mitochondrial membrane. PINK1 phosphorylates ubiquitin (Ub) and the ubiquitin ligase Parkin, and activated Parkin then ubiquitinates outer mitochondrial membrane proteins, leading to the recruitment of the autophagosome machinery and engulfment of the damaged organelle. Precise engulfment requires the Rab GAP TBC1D15 (shown in grey), which is bound to the mitochondrial outer membrane via interaction with LC3/GABARAP (not shown). The autophagosome then fuses with a lysosome, leading to degradation of the defective mitochondria by the proteases and lipases that reside in lysosomes.
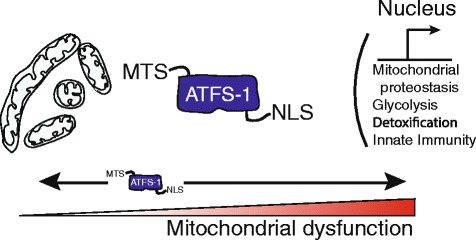
The ATFS-1-mediated mitochondrial unfolded protein response. The UPRmt is a protective transcriptional response to the accumulation of unfolded proteins or respiratory chain dysfunction that promotes adaptation and survival during mitochondrial dysfunction. Cells utilize the transcription factor ATFS-1 to monitor mitochondrial function and adjust transcription accordingly. Like PINK1, ATFS-1 is imported into mitochondria and quickly degraded in healthy cells, but accumulates in the cytosol during mitochondrial stress due to respiratory chain dysfunction, unfolded protein accumulation or high levels of reactive oxygen species, when mitochondrial import efficiency is impaired. Because ATFS-1 has a nuclear localization sequence (NLS), as well as its mitochondrial localization sequence (MTS), this causes it to localize to the nucleus, where it induces the transcription of genes involved in mitochondrial protein homeostasis, reactive oxygen species (ROS) and small molecule detoxification, glycolysis, and innate immunity.
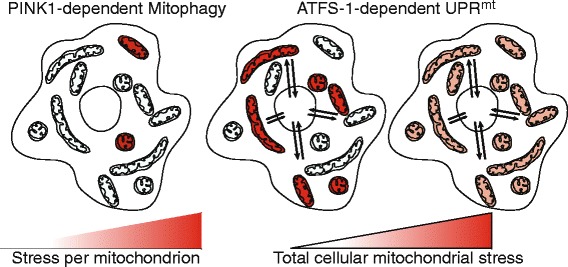
Integrating mitophagy and the UPR
mt
. While mitochondria are dynamic, at a given point in time they form a pool of at least semi-independent cellular compartments. Because PINK1 specifically accumulates on individual defective organelles (red), the mitophagy pathway is well equipped to monitor the health of individual organelles. On the other hand, ATFS-1 accumulates in the cytosol in proportion to the total amount of cellular mitochondrial stress. Because mitochondrial protein import is post-translational, ATFS-1 can accumulate in the cytosol during conditions where multiple individual mitochondria are severely defective (red; middle panel), or if the total pool of organelles is modestly stressed (far right panel). ATFS-1 induces transcription of a number of genes whose protein products must be imported into mitochondria to promote their recovery. Because the healthiest organelles are the most import competent, the UPRmt likely promotes the recovery of those organelles that can be salvaged. Concomitantly, by culling the most defective organelles, PINK1-dependent mitophagy enriches the healthy pool of organelles, allowing the resources required for mitochondrial repair to be allocated to the salvageable mitochondrial population.
Similar articles
-
Dysregulation of the Mitochondrial Unfolded Protein Response Induces Non-Apoptotic Dopaminergic Neurodegeneration in C. elegans Models of Parkinson's Disease.J Neurosci. 2017 Nov 15;37(46):11085-11100. doi: 10.1523/JNEUROSCI.1294-17.2017. Epub 2017 Oct 13. J Neurosci. 2017. PMID: 29030433 Free PMC article.
-
Mitophagy and neurodegeneration: the zebrafish model system.Autophagy. 2013 Nov 1;9(11):1693-709. doi: 10.4161/auto.25082. Epub 2013 Aug 8. Autophagy. 2013. PMID: 23939015 Review.
-
On the offense and defense: mitochondrial recovery programs amidst targeted pathogenic assault.FEBS J. 2022 Nov;289(22):7014-7037. doi: 10.1111/febs.16126. Epub 2021 Jul 30. FEBS J. 2022. PMID: 34270874 Free PMC article. Review.
-
The mitochondrial unfolded protein response and its diverse roles in cellular stress.Int J Biochem Cell Biol. 2021 Apr;133:105934. doi: 10.1016/j.biocel.2021.105934. Epub 2021 Jan 30. Int J Biochem Cell Biol. 2021. PMID: 33529716 Review.
-
The intersection of exercise and aging on mitochondrial protein quality control.Exp Gerontol. 2020 Mar;131:110824. doi: 10.1016/j.exger.2019.110824. Epub 2020 Jan 3. Exp Gerontol. 2020. PMID: 31911185 Review.
Cited by
-
Molecular Hydrogen Enhances Proliferation of Cancer Cells That Exhibit Potent Mitochondrial Unfolded Protein Response.Int J Mol Sci. 2022 Mar 7;23(5):2888. doi: 10.3390/ijms23052888. Int J Mol Sci. 2022. PMID: 35270030 Free PMC article.
-
Autophagic flux induced by graphene oxide has a neuroprotective effect against human prion protein fragments.Int J Nanomedicine. 2017 Nov 8;12:8143-8158. doi: 10.2147/IJN.S146398. eCollection 2017. Int J Nanomedicine. 2017. PMID: 29184404 Free PMC article.
-
A quantitative genome-wide RNAi screen in C. elegans for antifungal innate immunity genes.BMC Biol. 2016 Apr 29;14:35. doi: 10.1186/s12915-016-0256-3. BMC Biol. 2016. PMID: 27129311 Free PMC article.
-
Mitochondrial proteostasis in the context of cellular and organismal health and aging.J Biol Chem. 2019 Apr 5;294(14):5396-5407. doi: 10.1074/jbc.TM117.000893. Epub 2018 Apr 5. J Biol Chem. 2019. PMID: 29622680 Free PMC article. Review.
-
Interfacing mitochondrial biogenesis and elimination to enhance host pathogen defense and longevity.Worm. 2015 Jul 29;4(3):e1071763. doi: 10.1080/21624054.2015.1071763. eCollection 2015 Jul-Sep. Worm. 2015. PMID: 26430570 Free PMC article.
References
-
- van der Laan M, Hutu DP, Rehling P. On the mechanism of preprotein import by the mitochondrial presequence translocase. Biochim Biophys Acta. 1803;2010:732–9. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
