

ATR-mediated phosphorylation of FANCI regulates dormant origin firing in response to replication stress
- PMID: 25843623
- PMCID: PMC4408929
- DOI: 10.1016/j.molcel.2015.02.031
ATR-mediated phosphorylation of FANCI regulates dormant origin firing in response to replication stress
Abstract
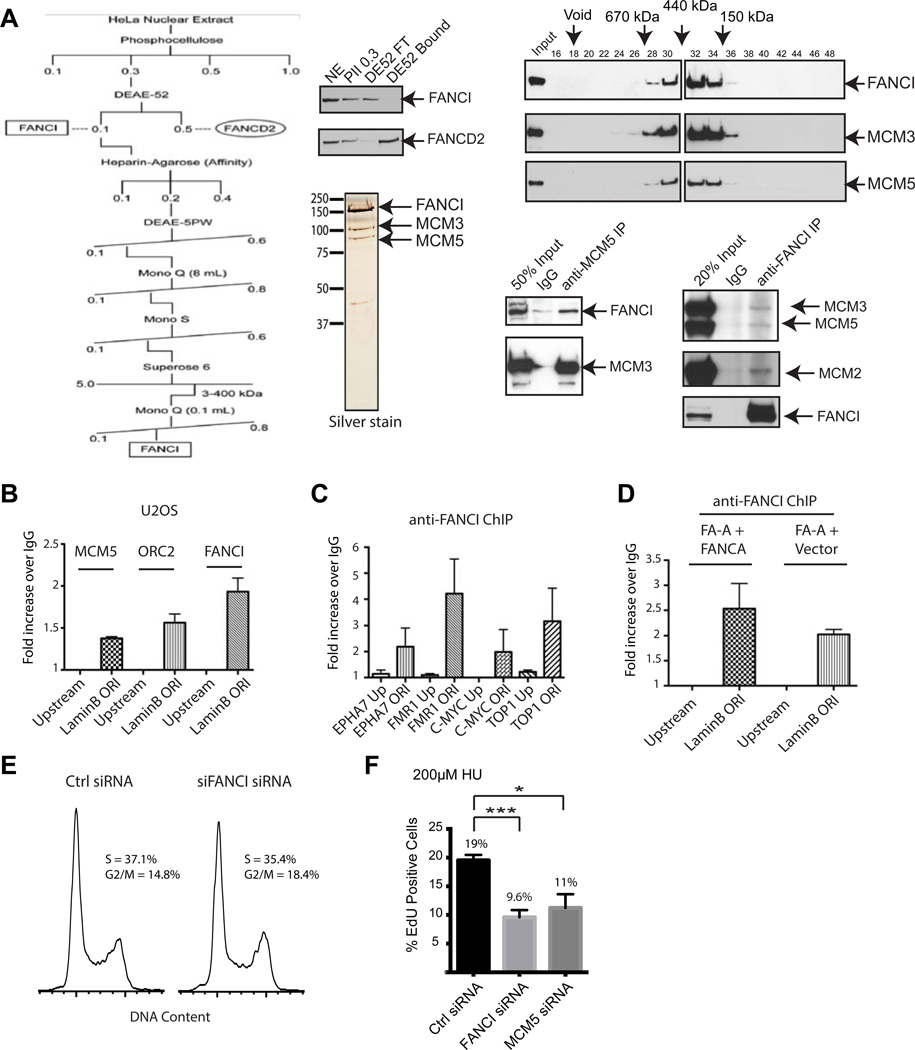
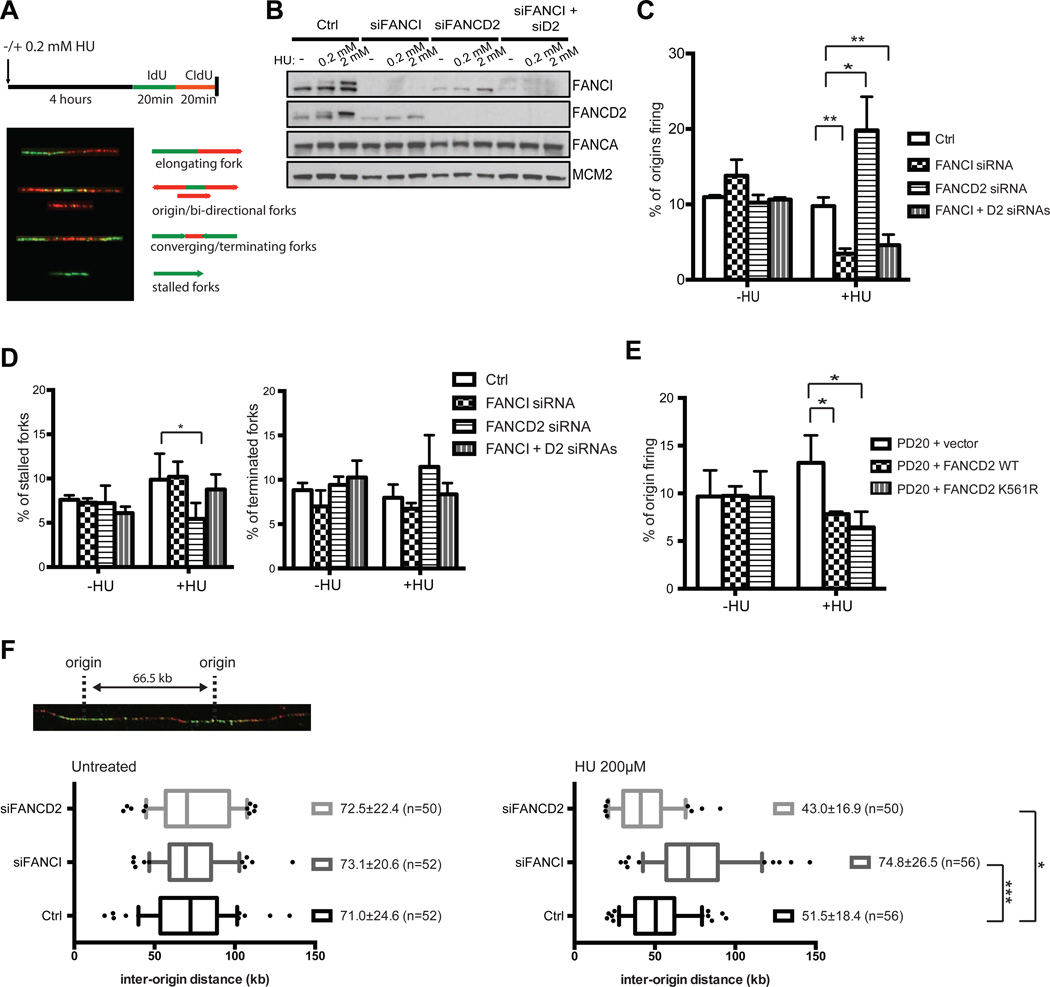
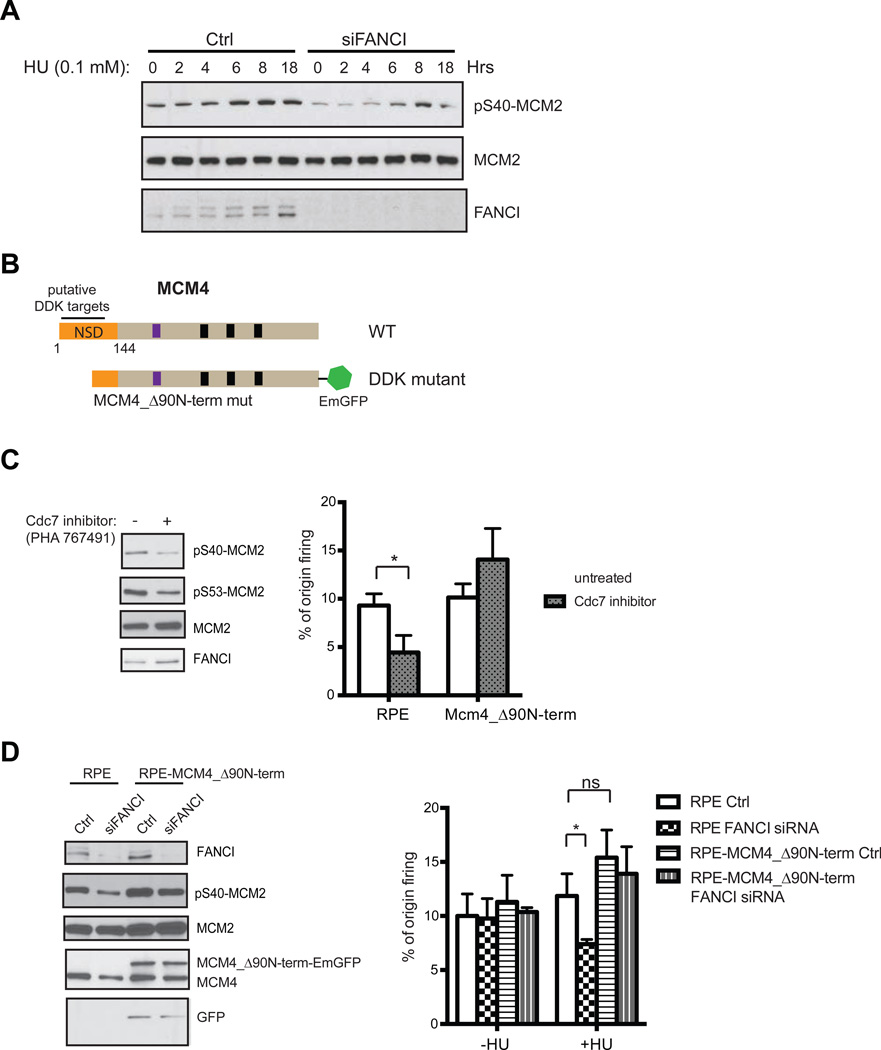
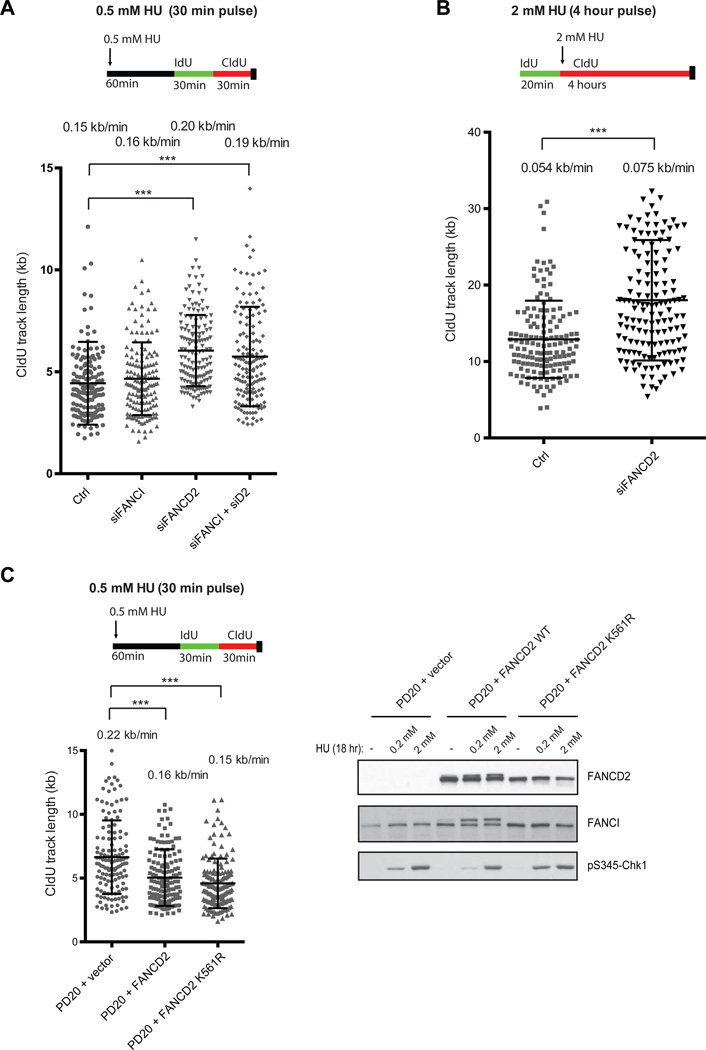
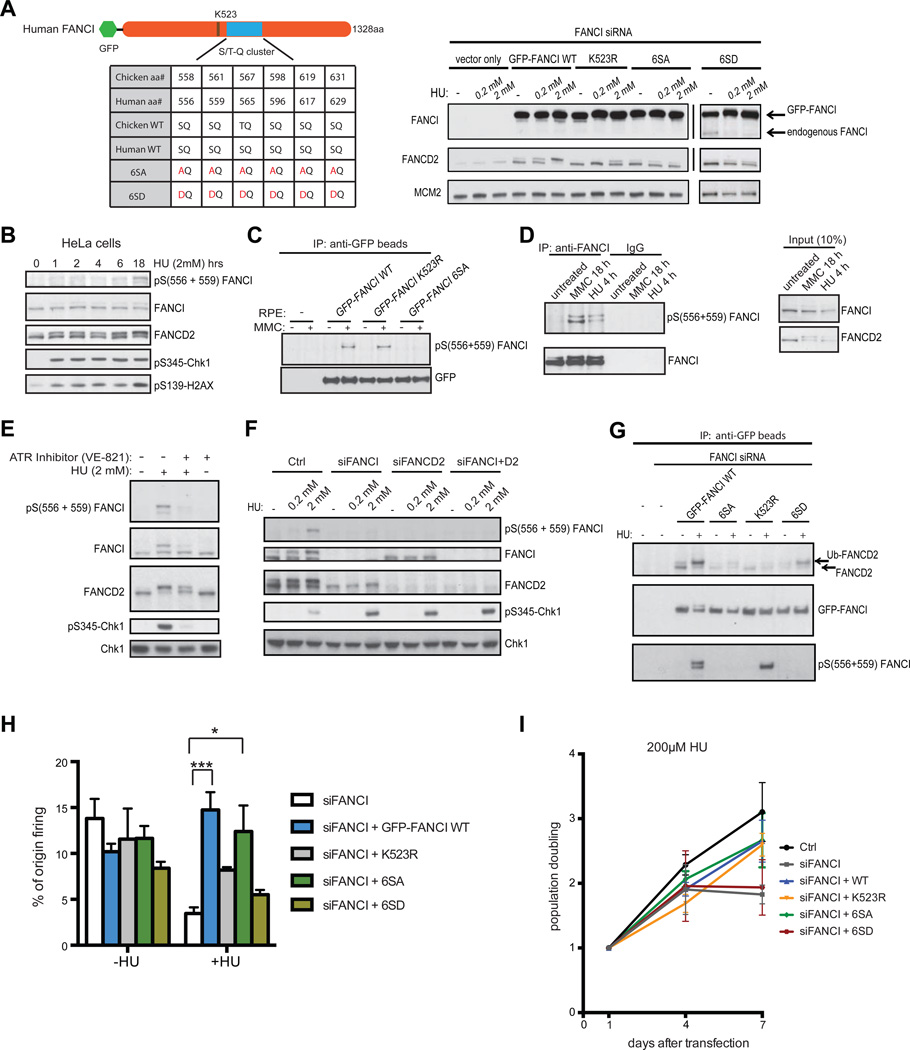
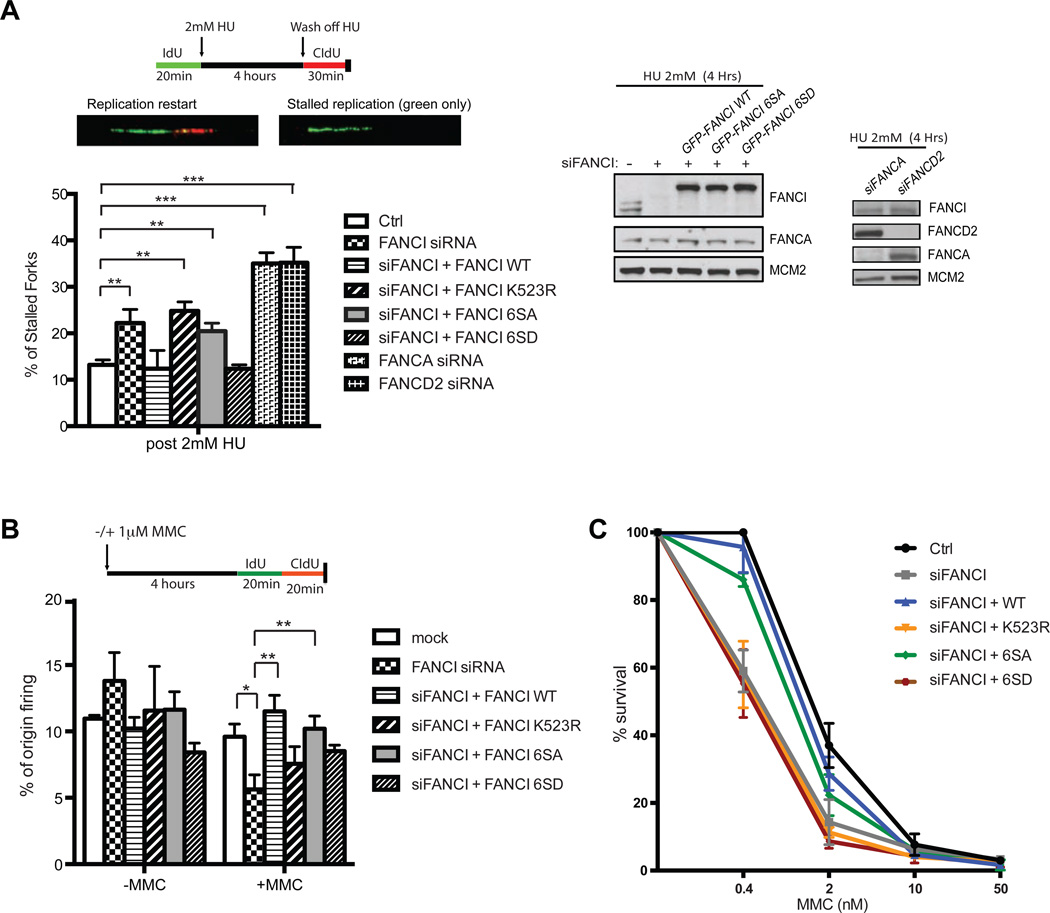
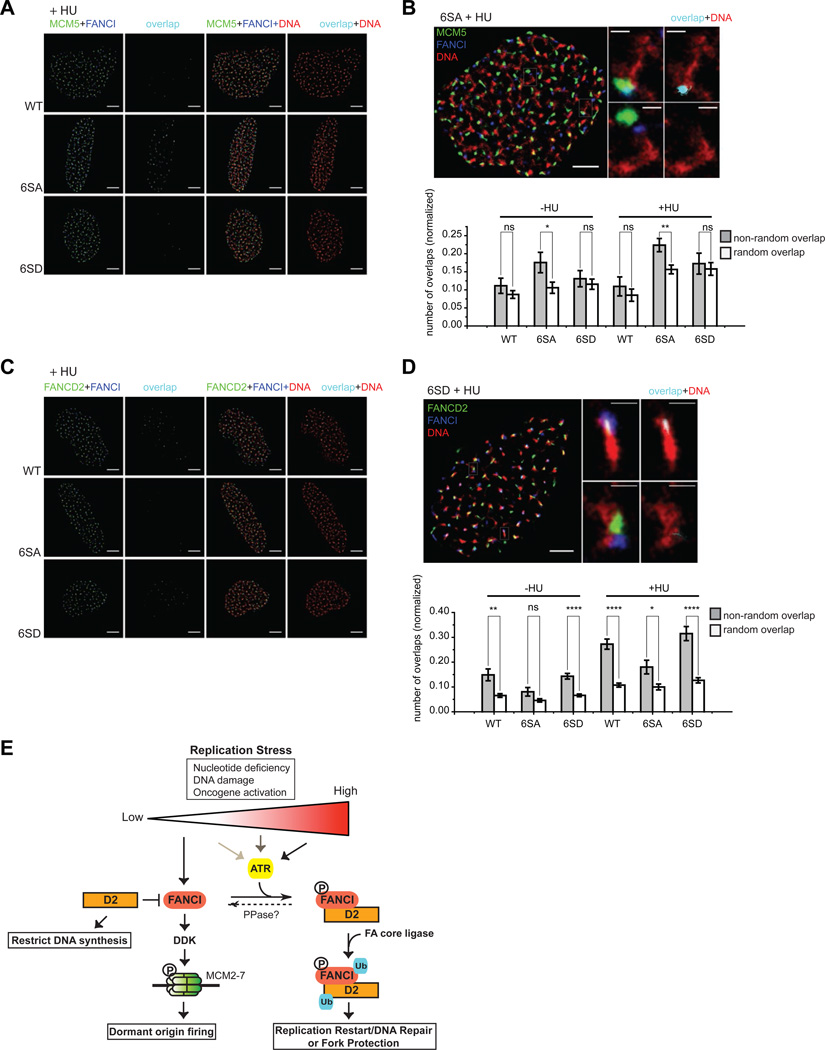
Excess dormant origins bound by the minichromosome maintenance (MCM) replicative helicase complex play a critical role in preventing replication stress, chromosome instability, and tumorigenesis. In response to DNA damage, replicating cells must coordinate DNA repair and dormant origin firing to ensure complete and timely replication of the genome; how cells regulate this process remains elusive. Herein, we identify a member of the Fanconi anemia (FA) DNA repair pathway, FANCI, as a key effector of dormant origin firing in response to replication stress. Cells lacking FANCI have reduced number of origins, increased inter-origin distances, and slowed proliferation rates. Intriguingly, ATR-mediated FANCI phosphorylation inhibits dormant origin firing while promoting replication fork restart/DNA repair. Using super-resolution microscopy, we show that FANCI co-localizes with MCM-bound chromatin in response to replication stress. These data reveal a unique role for FANCI as a modulator of dormant origin firing and link timely genome replication to DNA repair.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
FANCI phosphorylation functions as a molecular switch to turn on the Fanconi anemia pathway.Nat Struct Mol Biol. 2008 Nov;15(11):1138-46. doi: 10.1038/nsmb.1504. Epub 2008 Oct 19. Nat Struct Mol Biol. 2008. PMID: 18931676 Free PMC article.
-
FANCI and FANCD2 have common as well as independent functions during the cellular replication stress response.Nucleic Acids Res. 2017 Nov 16;45(20):11837-11857. doi: 10.1093/nar/gkx847. Nucleic Acids Res. 2017. PMID: 29059323 Free PMC article.
-
Fanconi anemia FANCD2 and FANCI proteins regulate the nuclear dynamics of splicing factors.J Cell Biol. 2017 Dec 4;216(12):4007-4026. doi: 10.1083/jcb.201702136. Epub 2017 Oct 13. J Cell Biol. 2017. PMID: 29030393 Free PMC article.
-
Behavior of replication origins in Eukaryota - spatio-temporal dynamics of licensing and firing.Cell Cycle. 2015;14(14):2251-64. doi: 10.1080/15384101.2015.1056421. Epub 2015 Jun 1. Cell Cycle. 2015. PMID: 26030591 Free PMC article. Review.
-
Dormant origin signaling during unperturbed replication.DNA Repair (Amst). 2019 Sep;81:102655. doi: 10.1016/j.dnarep.2019.102655. Epub 2019 Jul 8. DNA Repair (Amst). 2019. PMID: 31311769 Free PMC article. Review.
Cited by
-
Acetylation modulates the Fanconi anemia pathway by protecting FAAP20 from ubiquitin-mediated proteasomal degradation.J Biol Chem. 2020 Oct 2;295(40):13887-13901. doi: 10.1074/jbc.RA120.015288. Epub 2020 Aug 6. J Biol Chem. 2020. PMID: 32763975 Free PMC article.
-
Replication Protein A (RPA) deficiency activates the Fanconi anemia DNA repair pathway.Cell Cycle. 2016 Sep;15(17):2336-45. doi: 10.1080/15384101.2016.1201621. Epub 2016 Jul 11. Cell Cycle. 2016. PMID: 27398742 Free PMC article.
-
Spatiotemporal dynamics of homologous recombination repair at single collapsed replication forks.Nat Commun. 2018 Sep 24;9(1):3882. doi: 10.1038/s41467-018-06435-3. Nat Commun. 2018. PMID: 30250272 Free PMC article.
-
Molecular Genetic Characteristics of FANCI, a Proposed New Ovarian Cancer Predisposing Gene.Genes (Basel). 2023 Jan 20;14(2):277. doi: 10.3390/genes14020277. Genes (Basel). 2023. PMID: 36833203 Free PMC article.
-
Focal Point of Fanconi Anemia Signaling.Int J Mol Sci. 2021 Nov 30;22(23):12976. doi: 10.3390/ijms222312976. Int J Mol Sci. 2021. PMID: 34884777 Free PMC article. Review.
References
-
- Auerbach AD, Wolman SR. Susceptibility of Fanconi’s anaemia fibroblasts to chromosome damage by carcinogens. Nature. 1976;261:494–496. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
