

The Drosophila melanogaster Mutants apblot and apXasta Affect an Essential apterous Wing Enhancer
- PMID: 25840432
- PMCID: PMC4478543
- DOI: 10.1534/g3.115.017707
The Drosophila melanogaster Mutants apblot and apXasta Affect an Essential apterous Wing Enhancer
Abstract
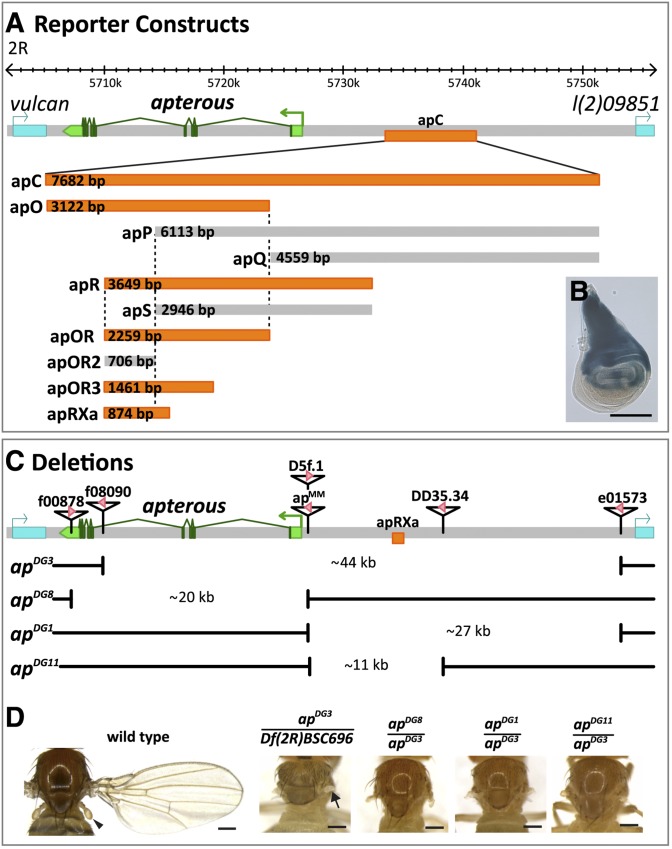
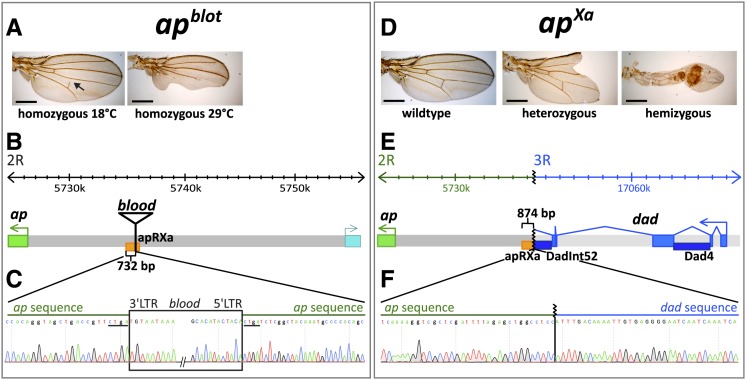
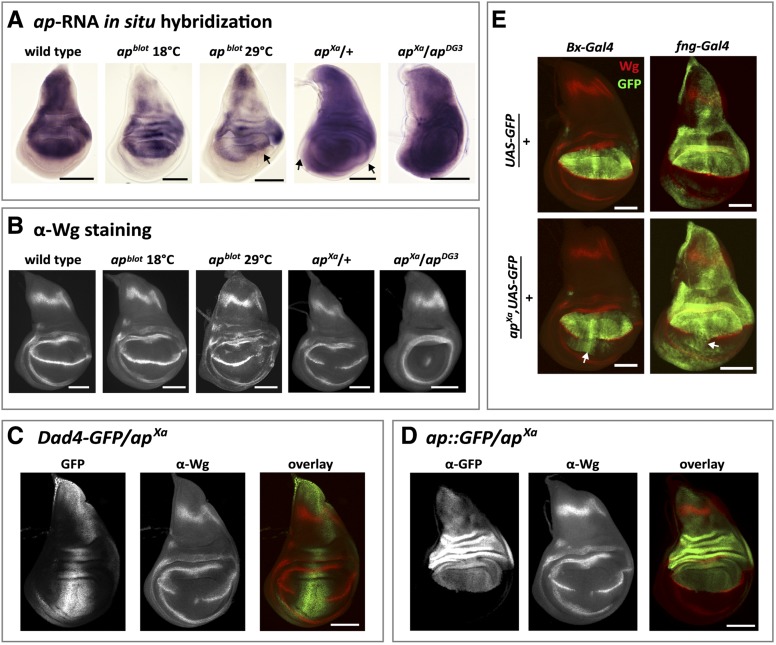
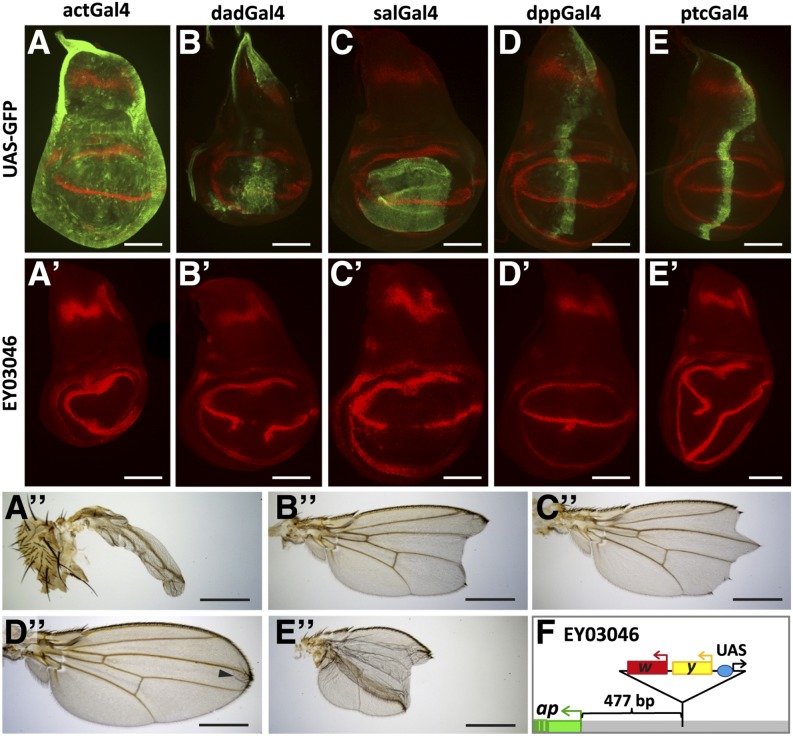
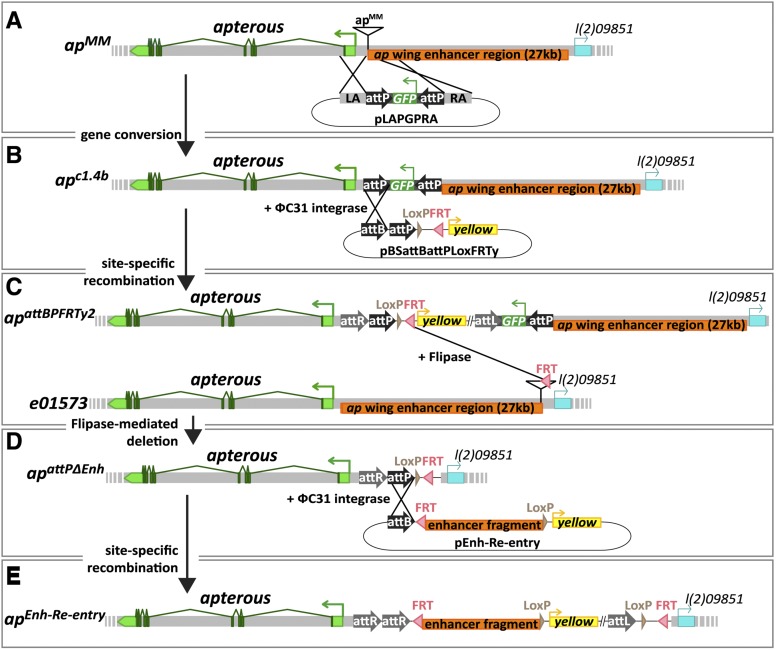
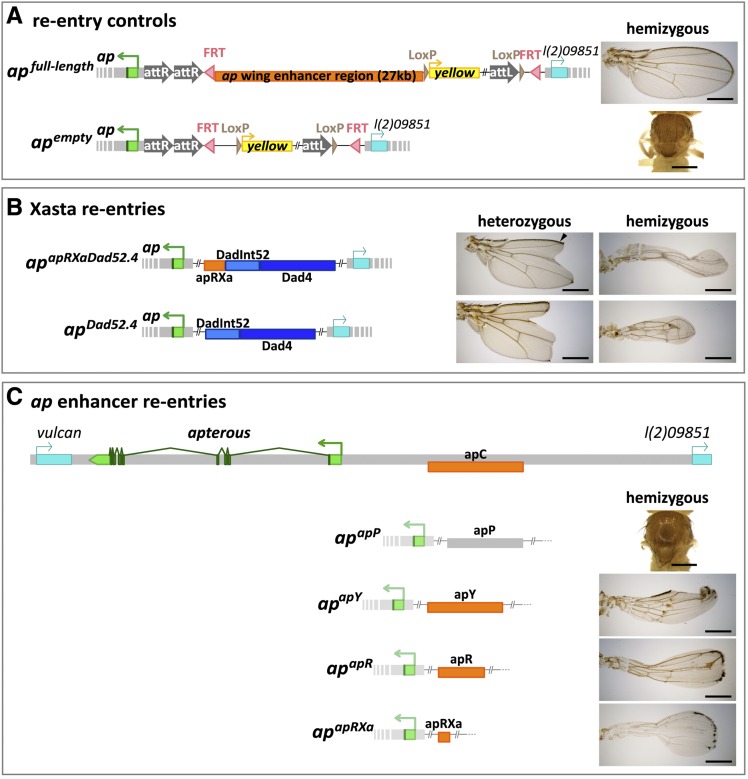
The selector gene apterous (ap) plays a key role during the development of the Drosophila melanogaster wing because it governs the establishment of the dorsal-ventral (D-V) compartment boundary. The D-V compartment boundary is known to serve as an important signaling center that is essential for the growth of the wing. The role of Ap and its downstream effectors have been studied extensively. However, very little is known about the transcriptional regulation of ap during wing disc development. In this study, we present a first characterization of an essential wing-specific ap enhancer. First, we defined an 874-bp fragment about 10 kb upstream of the ap transcription start that faithfully recapitulates the expression pattern of ap in the wing imaginal disc. Analysis of deletions in the ap locus covering this element demonstrated that it is essential for proper regulation of ap and formation of the wing. Moreover, we showed that the mutations ap(blot) and ap(Xasta) directly affect the integrity of this enhancer, leading to characteristic wing phenotypes. Furthermore, we engineered an in situ rescue system at the endogenous ap gene locus, allowing us to investigate the role of enhancer fragments in their native environment. Using this system, we were able to demonstrate that the essential wing enhancer alone is not sufficient for normal wing development. The in situ rescue system will allow us to characterize the ap regulatory sequences in great detail at the endogenous locus.
Keywords: Drosophila; apterous; boundary; compartment.
Copyright © 2015 Bieli et al.
Figures
Similar articles
-
Establishment of a Developmental Compartment Requires Interactions between Three Synergistic Cis-regulatory Modules.PLoS Genet. 2015 Oct 15;11(10):e1005376. doi: 10.1371/journal.pgen.1005376. eCollection 2015 Oct. PLoS Genet. 2015. PMID: 26468882 Free PMC article.
-
Enhancer blocking and transvection at the Drosophila apterous locus.Genetics. 2008 Jan;178(1):127-43. doi: 10.1534/genetics.107.077768. Genetics. 2008. PMID: 18202363 Free PMC article.
-
Drosophila wing development in the absence of dorsal identity.Development. 2001 Mar;128(5):703-10. doi: 10.1242/dev.128.5.703. Development. 2001. PMID: 11171395
-
Protocols to Study Growth and Metabolism in Drosophila.Methods Mol Biol. 2016;1478:279-290. doi: 10.1007/978-1-4939-6371-3_17. Methods Mol Biol. 2016. PMID: 27730589 Review.
-
Establishing compartment boundaries in Drosophila wing imaginal discs: An interplay between selector genes, signaling pathways and cell mechanics.Semin Cell Dev Biol. 2020 Nov;107:161-169. doi: 10.1016/j.semcdb.2020.07.008. Epub 2020 Jul 27. Semin Cell Dev Biol. 2020. PMID: 32732129 Review.
Cited by
-
Establishment of a Developmental Compartment Requires Interactions between Three Synergistic Cis-regulatory Modules.PLoS Genet. 2015 Oct 15;11(10):e1005376. doi: 10.1371/journal.pgen.1005376. eCollection 2015 Oct. PLoS Genet. 2015. PMID: 26468882 Free PMC article.
-
Neuronal cell fate diversification controlled by sub-temporal action of Kruppel.Elife. 2016 Oct 14;5:e19311. doi: 10.7554/eLife.19311. Elife. 2016. PMID: 27740908 Free PMC article.
-
Engineering the Drosophila Genome for Developmental Biology.J Dev Biol. 2017 Dec 11;5(4):16. doi: 10.3390/jdb5040016. J Dev Biol. 2017. PMID: 29615571 Free PMC article. Review.
-
Influence of genetic polymorphism on transcriptional enhancer activity in the malaria vector Anopheles coluzzii.Sci Rep. 2019 Oct 24;9(1):15275. doi: 10.1038/s41598-019-51730-8. Sci Rep. 2019. PMID: 31649293 Free PMC article.
-
Study on the Effect of Wing Bud Chitin Metabolism and Its Developmental Network Genes in the Brown Planthopper, Nilaparvata lugens, by Knockdown of TRE Gene.Front Physiol. 2017 Sep 26;8:750. doi: 10.3389/fphys.2017.00750. eCollection 2017. Front Physiol. 2017. PMID: 29033849 Free PMC article.
References
-
- Affolter M., Basler K., 2007. The decapentaplegic morphogen gradient: from pattern formation to growth regulation. Nat. Rev. Genet. 8: 663–674. - PubMed
-
- Ahmad K., Henikoff S., 2001. Modulation of a transcription factor counteracts heterochromatic gene silencing in Drosophila. Cell 104: 839–847. - PubMed
-
- Alexandre C., Jacinto A., Ingham P. W., 1996. Transcriptional activation of hedgehog target genes in Drosophila is mediated directly by the cubitus interruptus protein, a member of the GLI family of zinc finger DNA-binding proteins. Genes Dev. 10: 2003–2013. - PubMed
-
- Ashburner M., 1989. Drosophila: A Laboratory Handbook and Manual (two volumes) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases