

Biochemical characterization of a Naegleria TET-like oxygenase and its application in single molecule sequencing of 5-methylcytosine
- PMID: 25831492
- PMCID: PMC4394277
- DOI: 10.1073/pnas.1417939112
Biochemical characterization of a Naegleria TET-like oxygenase and its application in single molecule sequencing of 5-methylcytosine
Abstract
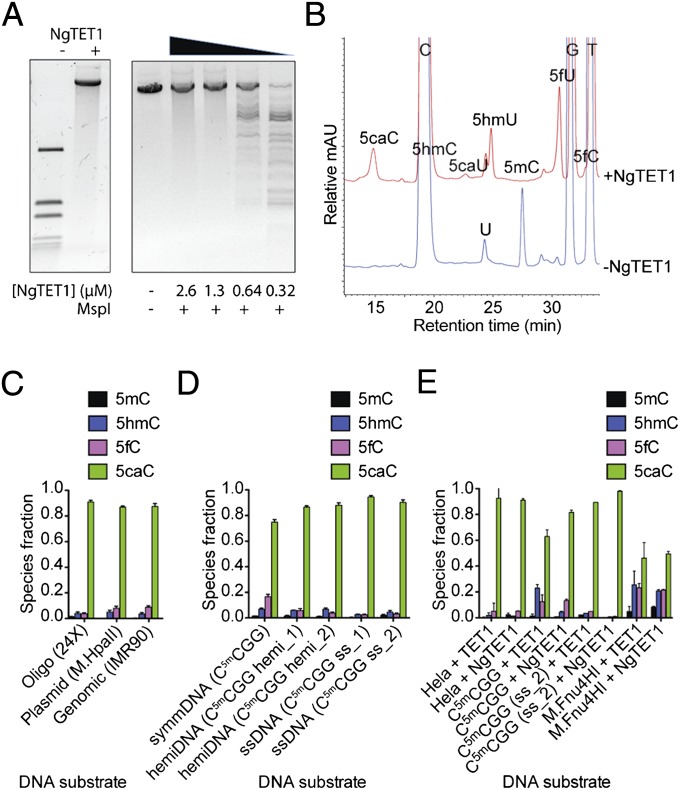
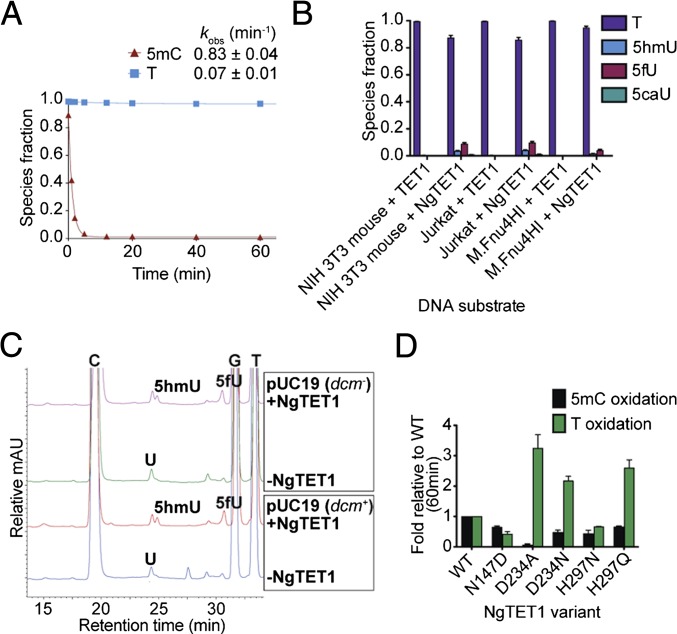
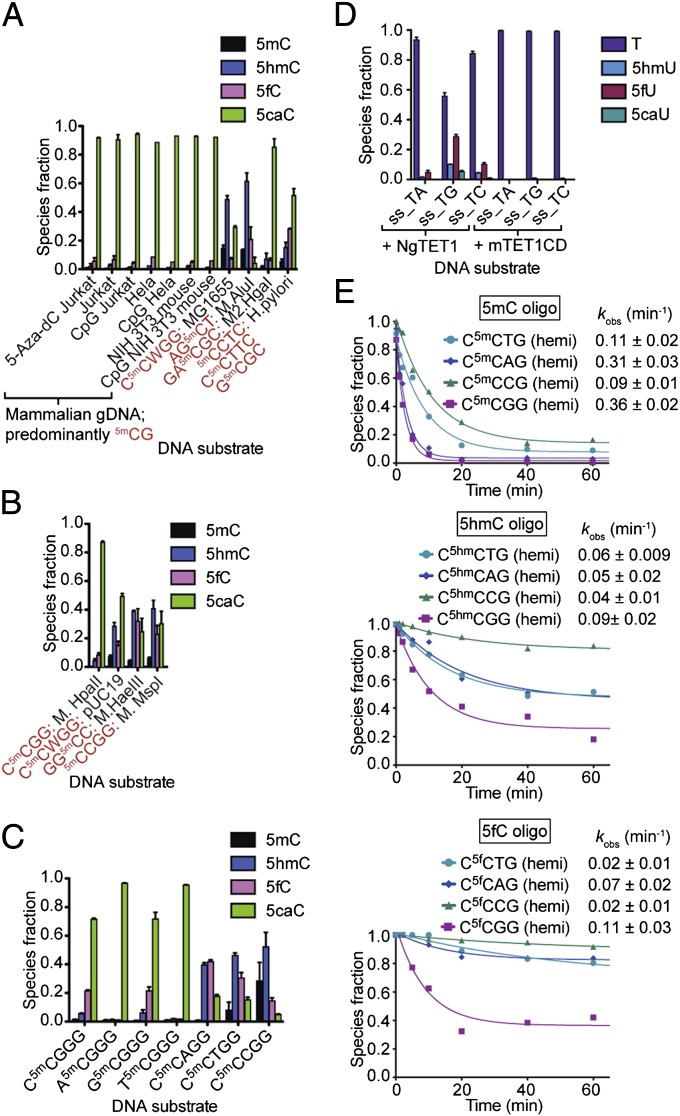
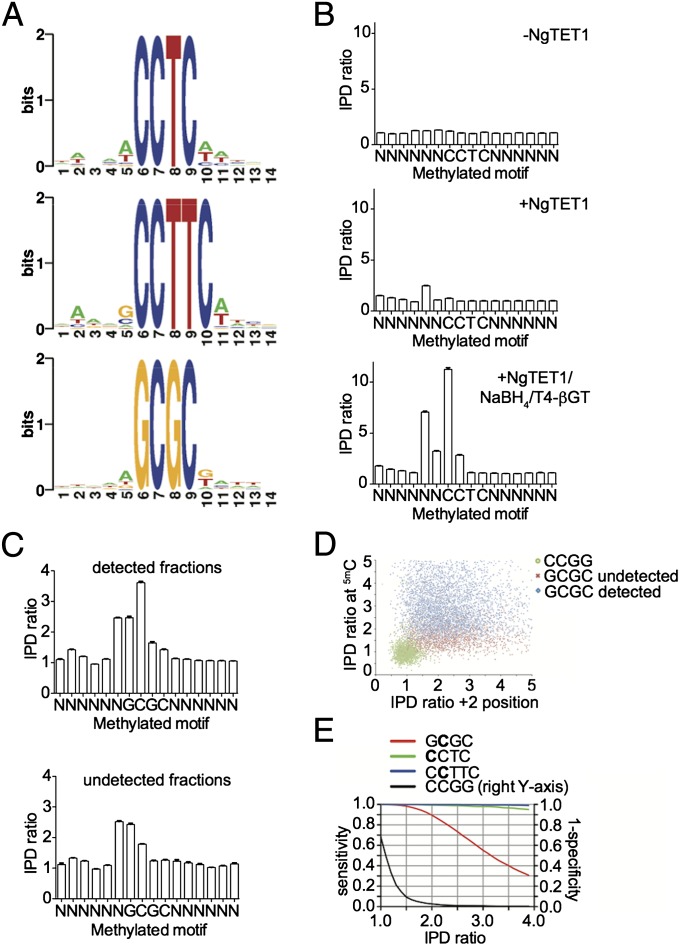
Modified DNA bases in mammalian genomes, such as 5-methylcytosine ((5m)C) and its oxidized forms, are implicated in important epigenetic regulation processes. In human or mouse, successive enzymatic conversion of (5m)C to its oxidized forms is carried out by the ten-eleven translocation (TET) proteins. Previously we reported the structure of a TET-like (5m)C oxygenase (NgTET1) from Naegleria gruberi, a single-celled protist evolutionarily distant from vertebrates. Here we show that NgTET1 is a 5-methylpyrimidine oxygenase, with activity on both (5m)C (major activity) and thymidine (T) (minor activity) in all DNA forms tested, and provide unprecedented evidence for the formation of 5-formyluridine ((5f)U) and 5-carboxyuridine ((5ca)U) in vitro. Mutagenesis studies reveal a delicate balance between choice of (5m)C or T as the preferred substrate. Furthermore, our results suggest substrate preference by NgTET1 to (5m)CpG and TpG dinucleotide sites in DNA. Intriguingly, NgTET1 displays higher T-oxidation activity in vitro than mammalian TET1, supporting a closer evolutionary relationship between NgTET1 and the base J-binding proteins from trypanosomes. Finally, we demonstrate that NgTET1 can be readily used as a tool in (5m)C sequencing technologies such as single molecule, real-time sequencing to map (5m)C in bacterial genomes at base resolution.
Keywords: 5-methylcytosine; NgTET1; SMRT sequencing; TET proteins; bacterial methylome.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Structure of a Naegleria Tet-like dioxygenase in complex with 5-methylcytosine DNA.Nature. 2014 Feb 20;506(7488):391-5. doi: 10.1038/nature12905. Epub 2013 Dec 25. Nature. 2014. PMID: 24390346 Free PMC article.
-
Distributive Processing by the Iron(II)/α-Ketoglutarate-Dependent Catalytic Domains of the TET Enzymes Is Consistent with Epigenetic Roles for Oxidized 5-Methylcytosine Bases.J Am Chem Soc. 2016 Aug 3;138(30):9345-8. doi: 10.1021/jacs.6b03243. Epub 2016 Jul 19. J Am Chem Soc. 2016. PMID: 27362828
-
Structure of Naegleria Tet-like dioxygenase (NgTet1) in complexes with a reaction intermediate 5-hydroxymethylcytosine DNA.Nucleic Acids Res. 2015 Dec 15;43(22):10713-21. doi: 10.1093/nar/gkv870. Epub 2015 Aug 31. Nucleic Acids Res. 2015. PMID: 26323320 Free PMC article.
-
TET enzymatic oxidation of 5-methylcytosine, 5-hydroxymethylcytosine and 5-formylcytosine.Mutat Res Genet Toxicol Environ Mutagen. 2014 Apr;764-765:18-35. doi: 10.1016/j.mrgentox.2013.09.001. Epub 2013 Sep 14. Mutat Res Genet Toxicol Environ Mutagen. 2014. PMID: 24045206 Review.
-
5-Methylcytosine DNA demethylation: more than losing a methyl group.Annu Rev Genet. 2012;46:419-41. doi: 10.1146/annurev-genet-110711-155451. Epub 2012 Sep 4. Annu Rev Genet. 2012. PMID: 22974304 Review.
Cited by
-
TET-TDG Active DNA Demethylation at CpG and Non-CpG Sites.J Mol Biol. 2021 Apr 16;433(8):166877. doi: 10.1016/j.jmb.2021.166877. Epub 2021 Feb 7. J Mol Biol. 2021. PMID: 33561435 Free PMC article.
-
Recent Advances on DNA Base Flipping: A General Mechanism for Writing, Reading, and Erasing DNA Modifications.Adv Exp Med Biol. 2022;1389:295-315. doi: 10.1007/978-3-031-11454-0_12. Adv Exp Med Biol. 2022. PMID: 36350515 Free PMC article.
-
Synthesis and biophysical analysis of modified thymine-containing DNA oligonucleotides.Chem Commun (Camb). 2017 Jan 24;53(8):1389-1392. doi: 10.1039/c6cc08670e. Chem Commun (Camb). 2017. PMID: 28074944 Free PMC article.
-
Dynamic changes in genomic 5-hydroxymethyluracil and N6-methyladenine levels in the Drosophila melanogaster life cycle and in response to different temperature conditions.Sci Rep. 2022 Oct 20;12(1):17552. doi: 10.1038/s41598-022-22490-9. Sci Rep. 2022. PMID: 36266436 Free PMC article.
-
Distinctive Klf4 mutants determine preference for DNA methylation status.Nucleic Acids Res. 2016 Dec 1;44(21):10177-10185. doi: 10.1093/nar/gkw774. Epub 2016 Sep 4. Nucleic Acids Res. 2016. PMID: 27596594 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
