

Co-chaperone p23 regulates C. elegans Lifespan in Response to Temperature
- PMID: 25830239
- PMCID: PMC4382338
- DOI: 10.1371/journal.pgen.1005023
Co-chaperone p23 regulates C. elegans Lifespan in Response to Temperature
Abstract
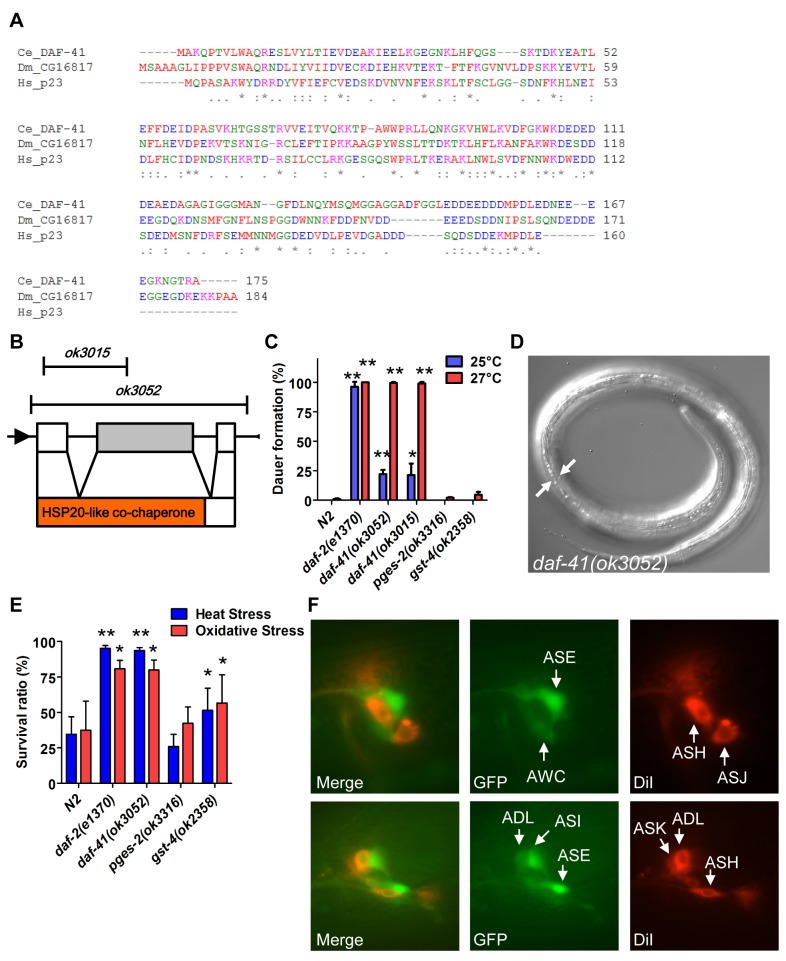
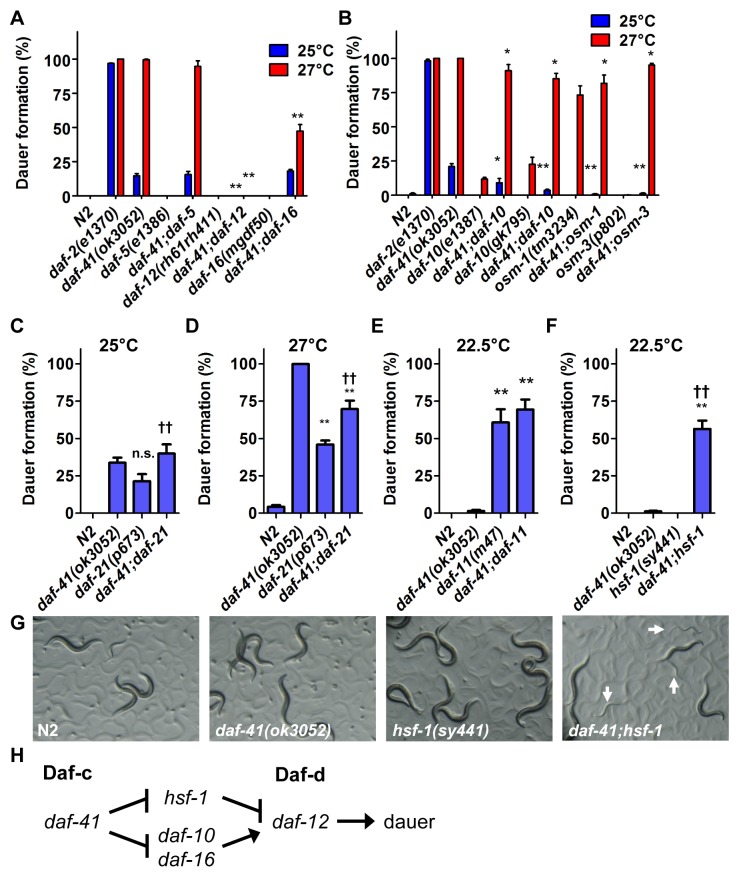
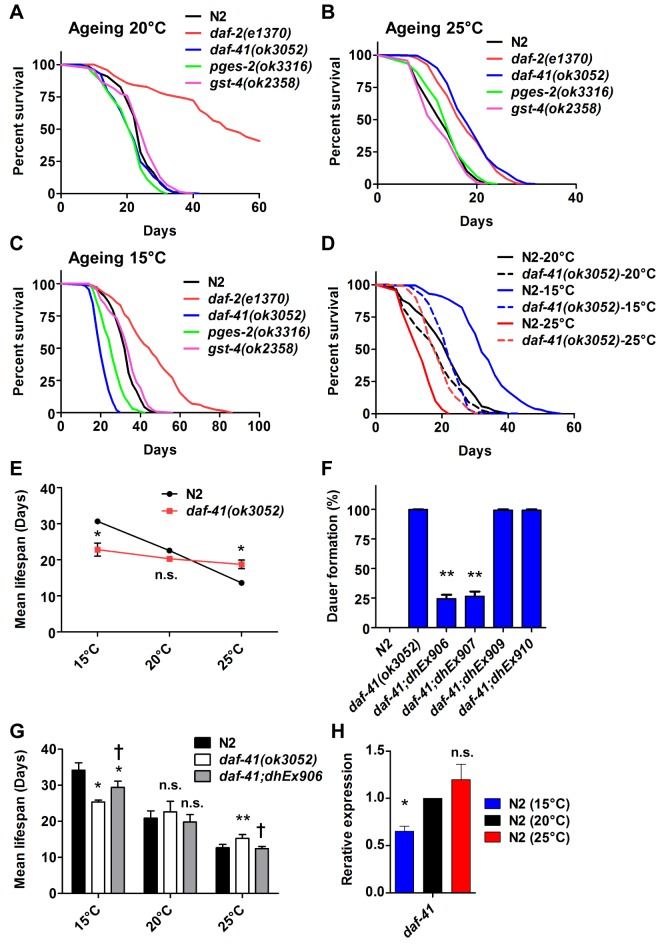
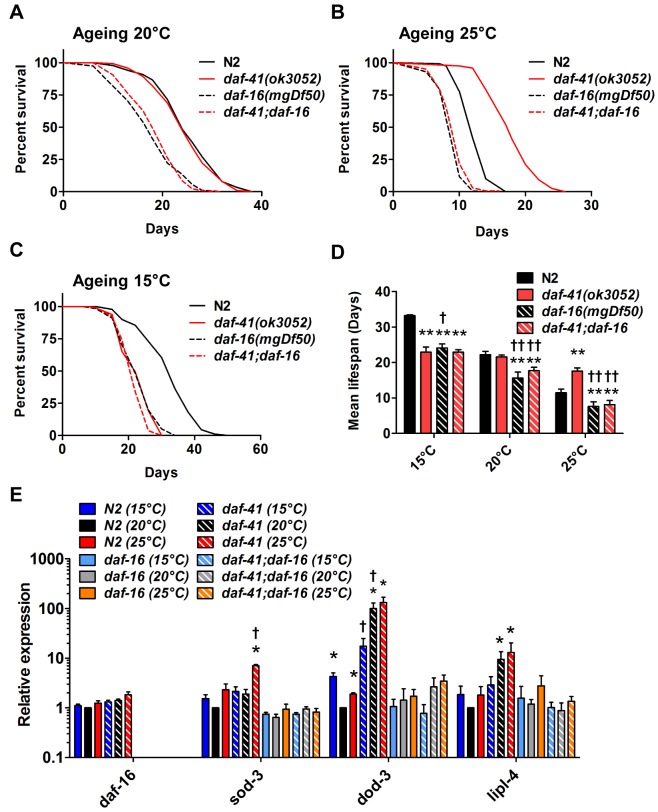
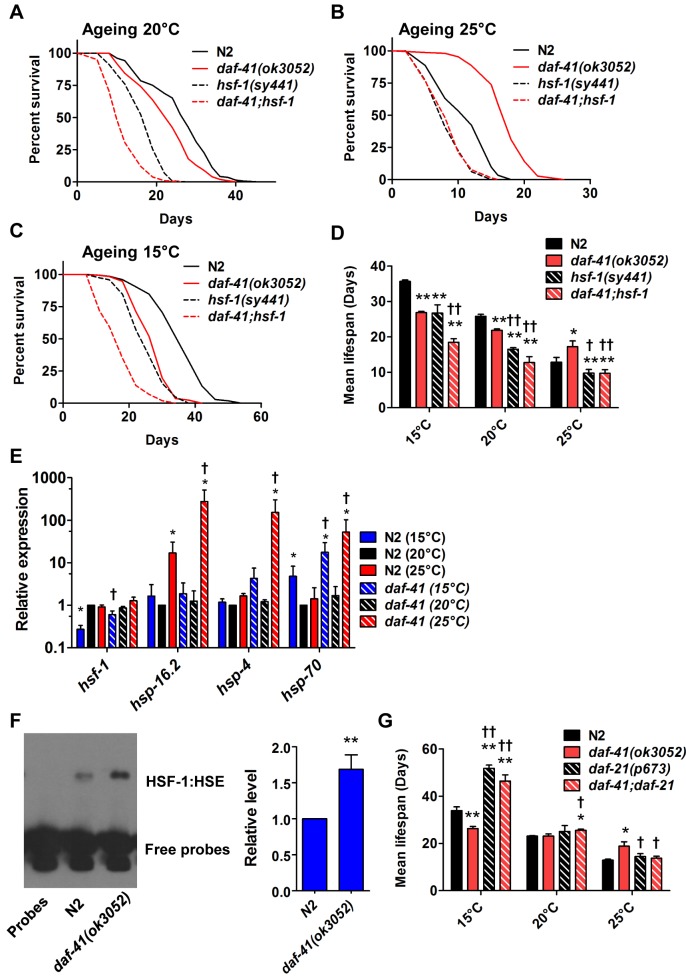
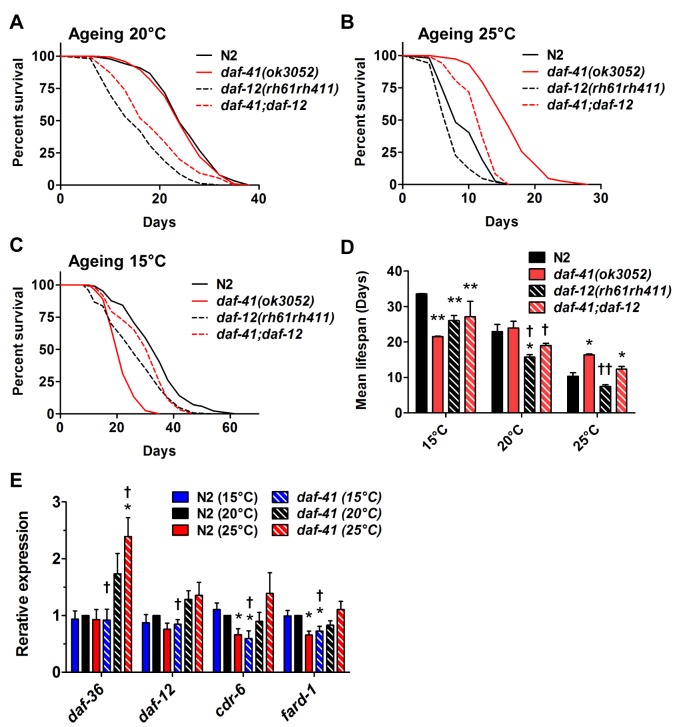
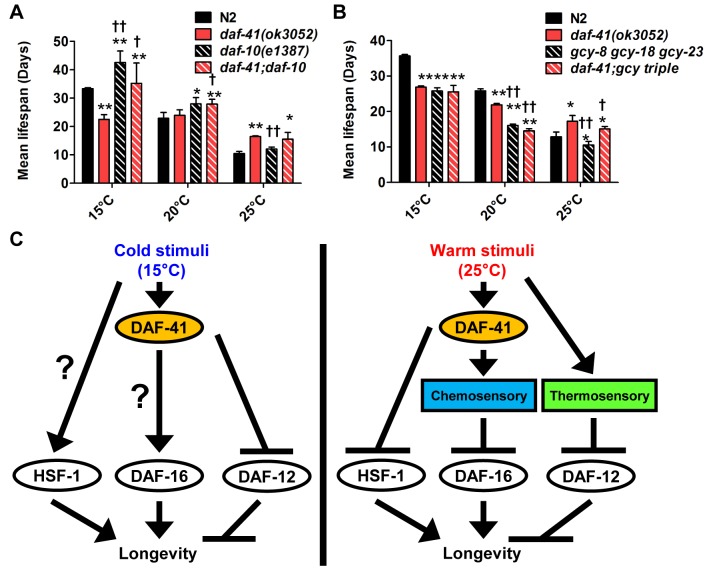
Temperature potently modulates various physiologic processes including organismal motility, growth rate, reproduction, and ageing. In ectotherms, longevity varies inversely with temperature, with animals living shorter at higher temperatures. Thermal effects on lifespan and other processes are ascribed to passive changes in metabolic rate, but recent evidence also suggests a regulated process. Here, we demonstrate that in response to temperature, daf-41/ZC395.10, the C. elegans homolog of p23 co-chaperone/prostaglandin E synthase-3, governs entry into the long-lived dauer diapause and regulates adult lifespan. daf-41 deletion triggers constitutive entry into the dauer diapause at elevated temperature dependent on neurosensory machinery (daf-10/IFT122), insulin/IGF-1 signaling (daf-16/FOXO), and steroidal signaling (daf-12/FXR). Surprisingly, daf-41 mutation alters the longevity response to temperature, living longer than wild-type at 25°C but shorter than wild-type at 15°C. Longevity phenotypes at 25°C work through daf-16/FOXO and heat shock factor hsf-1, while short lived phenotypes converge on daf-16/FOXO and depend on the daf-12/FXR steroid receptor. Correlatively daf-41 affected expression of DAF-16 and HSF-1 target genes at high temperature, and nuclear extracts from daf-41 animals showed increased occupancy of the heat shock response element. Our studies suggest that daf-41/p23 modulates key transcriptional changes in longevity pathways in response to temperature.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Caenorhabditis elegans EAK-3 inhibits dauer arrest via nonautonomous regulation of nuclear DAF-16/FoxO activity.Dev Biol. 2008 Mar 15;315(2):290-302. doi: 10.1016/j.ydbio.2007.12.032. Epub 2008 Jan 3. Dev Biol. 2008. PMID: 18241854 Free PMC article.
-
Prefoldin 6 mediates longevity response from heat shock factor 1 to FOXO in C. elegans.Genes Dev. 2018 Dec 1;32(23-24):1562-1575. doi: 10.1101/gad.317362.118. Epub 2018 Nov 26. Genes Dev. 2018. PMID: 30478249 Free PMC article.
-
DAF-21/Hsp90 is required for C. elegans longevity by ensuring DAF-16/FOXO isoform A function.Sci Rep. 2018 Aug 13;8(1):12048. doi: 10.1038/s41598-018-30592-6. Sci Rep. 2018. PMID: 30104664 Free PMC article.
-
DAF-16: FOXO in the Context of C. elegans.Curr Top Dev Biol. 2018;127:1-21. doi: 10.1016/bs.ctdb.2017.11.007. Epub 2018 Feb 2. Curr Top Dev Biol. 2018. PMID: 29433733 Review.
-
Worming pathways to and from DAF-16/FOXO.Exp Gerontol. 2006 Oct;41(10):928-34. doi: 10.1016/j.exger.2006.05.020. Epub 2006 Jul 12. Exp Gerontol. 2006. PMID: 16839734 Review.
Cited by
-
Regulatory mechanism of cold-inducible diapause in Caenorhabditis elegans.Nat Commun. 2024 Jul 10;15(1):5793. doi: 10.1038/s41467-024-50111-8. Nat Commun. 2024. PMID: 38987256 Free PMC article.
-
Prostaglandin signals from adult germ stem cells delay somatic aging of Caenorhabditis elegans.Nat Metab. 2019 Aug;1(8):790-810. doi: 10.1038/s42255-019-0097-9. Epub 2019 Aug 19. Nat Metab. 2019. PMID: 31485561 Free PMC article.
-
Screening for cold tolerance genes in C. elegans, whose expressions are affected by anticancer drugs camptothecin and leptomycin B.Sci Rep. 2024 Mar 5;14(1):5401. doi: 10.1038/s41598-024-55794-z. Sci Rep. 2024. PMID: 38443452 Free PMC article.
-
Caenorhabditis elegans orthologs of human genes differentially expressed with age are enriched for determinants of longevity.Aging Cell. 2017 Aug;16(4):672-682. doi: 10.1111/acel.12595. Epub 2017 Apr 12. Aging Cell. 2017. PMID: 28401650 Free PMC article.
-
Dietary restriction induces posttranscriptional regulation of longevity genes.Life Sci Alliance. 2019 Jun 28;2(4):e201800281. doi: 10.26508/lsa.201800281. Print 2019 Aug. Life Sci Alliance. 2019. PMID: 31253655 Free PMC article.
References
-
- Miquel J, Lundgren PR, Bensch KG, Atlan H (1976) Effects of temperature on the life span, vitality and fine structure of Drosophila melanogaster. Mech Ageing Dev 5: 347–370. - PubMed
-
- Klass MR (1977) Aging in the nematode Caenorhabditis elegans: major biological and environmental factors influencing life span. Mech Ageing Dev 6: 413–429. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
