

The RNA recognition motif of NIFK is required for rRNA maturation during cell cycle progression
- PMID: 25826659
- PMCID: PMC4615200
- DOI: 10.1080/15476286.2015.1017221
The RNA recognition motif of NIFK is required for rRNA maturation during cell cycle progression
Abstract
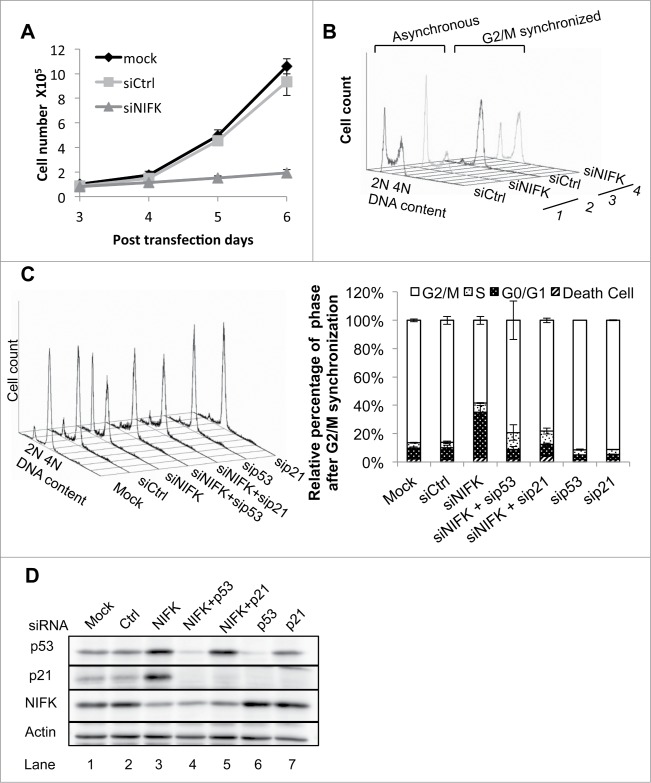
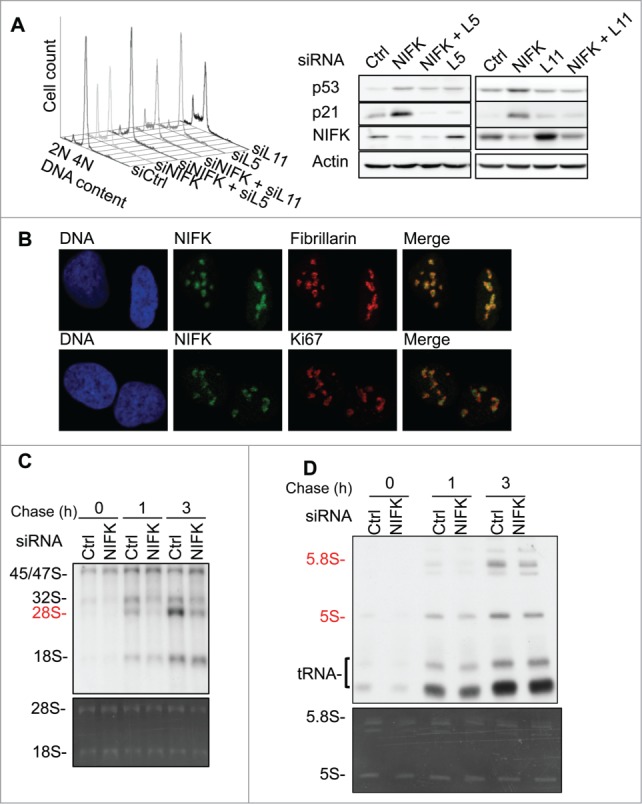
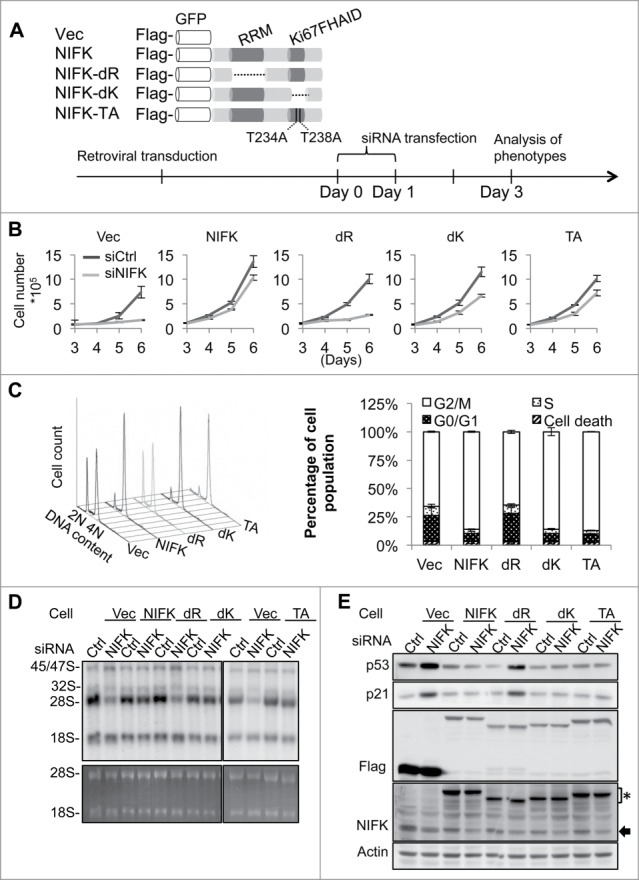
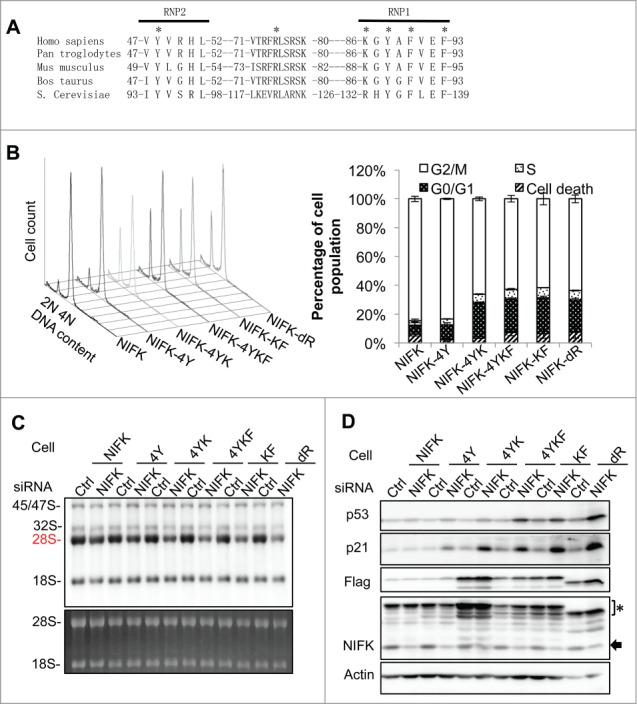
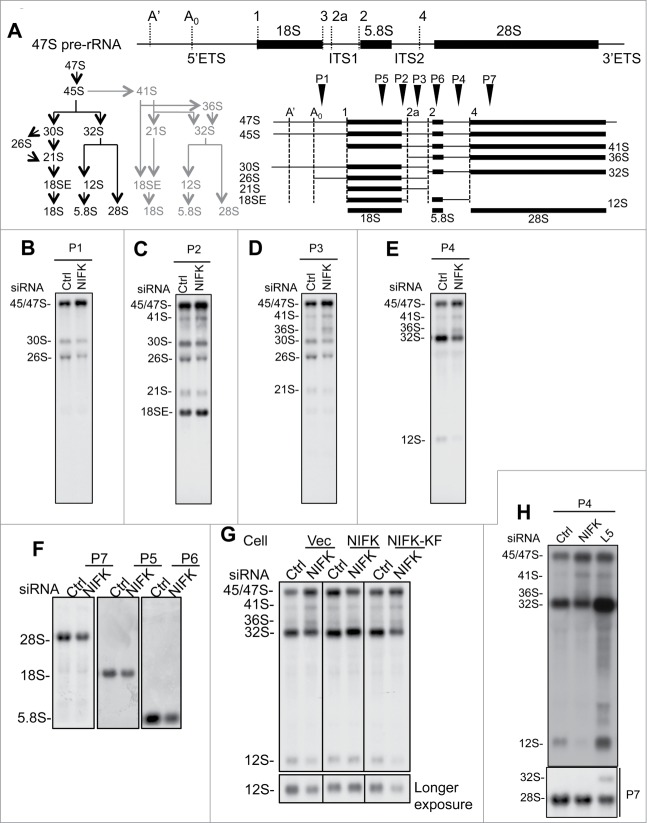
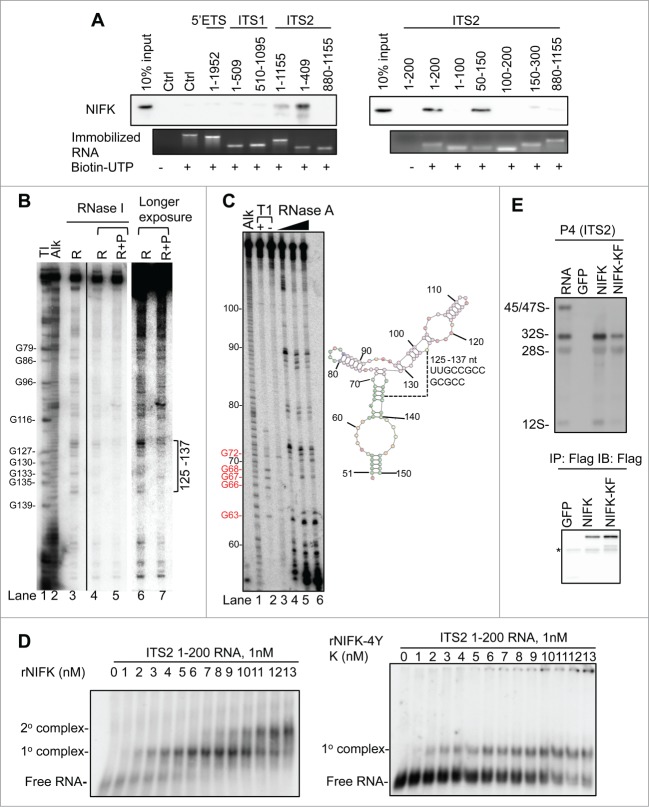
Ribosome biogenesis governs protein synthesis. NIFK is transactivated by c-Myc, the key regulator of ribosome biogenesis. The biological function of human NIFK is not well established, except that it has been shown to interact with Ki67 and NPM1. Here we report that NIFK is required for cell cycle progression and participates in the ribosome biogenesis via its RNA recognition motif (RRM). We show that silencing of NIFK inhibits cell proliferation through a reversible p53-dependent G1 arrest, possibly by induction of the RPL5/RPL11-mediated nucleolar stress. Mechanistically it is the consequence of impaired maturation of 28S and 5.8S rRNA resulting from inefficient cleavage of internal transcribed spacer (ITS) 1, a critical step in the separation of pre-ribosome to small and large subunits. Complementation of NIFK silencing by mutants shows that RNA-binding ability of RRM is essential for the pre-rRNA processing and G1 progression. More specifically, we validate that the RRM of NIFK preferentially binds to the 5'-region of ITS2 rRNA likely in both sequence specific and secondary structure dependent manners. Our results show how NIFK is involved in cell cycle progression through RRM-dependent pre-rRNA maturation, which could enhance our understanding of the function of NIFK in cell proliferation, and potentially also cancer and ribosomopathies.
Keywords: 5S RNP, 5S ribonucleoprotein particle; CDK1, cyclin dependent kinase 1; DFC, dense fibrillar component; ETS/ITS, external/internal transcribed spacers; GSK3, glycogen synthase kinase 3; Ki67; Ki67FHAID, Ki67-FHA interaction domain; LSU, large subunit; MDM2, murine double minute 2; NIFK, Nucleolar protein Interacting with the FHA domain of pKi-67; NPM1/B23, nucleophosmin; Noprecipitation; PAR-CLIP, Photo-Activatable-Ribonucleoside-Enhanced Crosslinking and Immu-pre-rRNAs, rRNA precursors; REMSA, RNA electrophoresis mobility shift assay; RNA recognition motif; RNP1 and 2, ribonucleoprotein motif 1 and 2; RPL5 and RPL11, large ribosomal protein 5 and 11; RRM, RNA recognition motif; cell cycle; nucleolar stress; rNIFK, recombinant NIFK; ribosome biogenesis; snoRNP, small nucleolar ribonucleoprotein.
Figures
Similar articles
-
The DExD box ATPase DDX55 is recruited to domain IV of the 28S ribosomal RNA by its C-terminal region.RNA Biol. 2021 Aug;18(8):1124-1135. doi: 10.1080/15476286.2020.1829366. Epub 2020 Oct 13. RNA Biol. 2021. PMID: 33048000 Free PMC article.
-
Bop1 is a mouse WD40 repeat nucleolar protein involved in 28S and 5. 8S RRNA processing and 60S ribosome biogenesis.Mol Cell Biol. 2000 Aug;20(15):5516-28. doi: 10.1128/MCB.20.15.5516-5528.2000. Mol Cell Biol. 2000. PMID: 10891491 Free PMC article.
-
Critical role of nucleostemin in pre-rRNA processing.J Biol Chem. 2009 Feb 20;284(8):4968-77. doi: 10.1074/jbc.M804594200. Epub 2008 Dec 23. J Biol Chem. 2009. PMID: 19106111 Free PMC article.
-
The multifunctional nucleolus.Nat Rev Mol Cell Biol. 2007 Jul;8(7):574-85. doi: 10.1038/nrm2184. Nat Rev Mol Cell Biol. 2007. PMID: 17519961 Review.
-
Activation of the tumor suppressor p53 upon impairment of ribosome biogenesis.Biochim Biophys Acta. 2014 Jun;1842(6):817-30. doi: 10.1016/j.bbadis.2013.08.014. Epub 2013 Oct 26. Biochim Biophys Acta. 2014. PMID: 24514102 Review.
Cited by
-
New insights into the biology of acute myeloid leukemia with mutated NPM1.Int J Hematol. 2019 Aug;110(2):150-160. doi: 10.1007/s12185-018-02578-7. Epub 2019 Jan 10. Int J Hematol. 2019. PMID: 30632059 Review.
-
MCRS1 Expression Regulates Tumor Activity and Affects Survival Probability of Patients with Gastric Cancer.Diagnostics (Basel). 2022 Jun 20;12(6):1502. doi: 10.3390/diagnostics12061502. Diagnostics (Basel). 2022. PMID: 35741311 Free PMC article.
-
Upregulation of GTPBP4 Promotes the Proliferation of Liver Cancer Cells.J Oncol. 2021 Oct 19;2021:1049104. doi: 10.1155/2021/1049104. eCollection 2021. J Oncol. 2021. PMID: 34712323 Free PMC article.
-
The Caenorhabditis elegans anchor cell transcriptome: ribosome biogenesis drives cell invasion through basement membrane.Development. 2023 May 1;150(9):dev201570. doi: 10.1242/dev.201570. Epub 2023 May 4. Development. 2023. PMID: 37039075 Free PMC article.
-
Identification of Differently Expressed Genes Associated With Prognosis and Growth in Colon Adenocarcinoma Based on Integrated Bioinformatics Analysis.Front Genet. 2019 Dec 4;10:1245. doi: 10.3389/fgene.2019.01245. eCollection 2019. Front Genet. 2019. PMID: 31867042 Free PMC article.
References
-
- Gerdes J, Lemke H, Baisch H, Wacker HH, Schwab U, Stein H. Cell cycle analysis of a cell proliferation-associated human nuclear antigen defined by the monoclonal antibody Ki-67. J Immunol 1984; 133:1710–5; PMID:6206131 - PubMed
-
- Scholzen T, Gerdes J. The Ki-67 protein: from the known and the unknown. J Cellular Physiol 2000; 182:311–22; PMID:10653597; http://dx.doi.org/10.1002/(SICI)1097-4652(200003)182:3%3c311::AID-JCP1%3... - DOI - PubMed
-
- Takagi M, Sueishi M, Saiwaki T, Kametaka A, Yoneda Y. A novel nucleolar protein, NIFK, interacts with the forkhead associated domain of Ki-67 antigen in mitosis. J Biol Chem 2001; 276:25386–91; PMID:11342549; http://dx.doi.org/10.1074/jbc.M102227200 - DOI - PubMed
-
- Byeon IJ, Li H, Song H, Gronenborn AM, Tsai MD. Sequential phosphorylation and multisite interactions characterize specific target recognition by the FHA domain of Ki67. Nat Struct Mol Biol 2005; 12:987–93; PMID:16244663; http://dx.doi.org/10.1038/nsmb1008 - DOI - PubMed
-
- Schlosser I, Holzel M, Murnseer M, Burtscher H, Weidle UH, Eick D. A role for c-Myc in the regulation of ribosomal RNA processing. Nucleic Acids Res 2003; 31:6148–56; PMID:14576301; http://dx.doi.org/10.1093/nar/gkg794 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous