

Osteopetrorickets due to Snx10 deficiency in mice results from both failed osteoclast activity and loss of gastric acid-dependent calcium absorption
- PMID: 25811986
- PMCID: PMC4374855
- DOI: 10.1371/journal.pgen.1005057
Osteopetrorickets due to Snx10 deficiency in mice results from both failed osteoclast activity and loss of gastric acid-dependent calcium absorption
Abstract
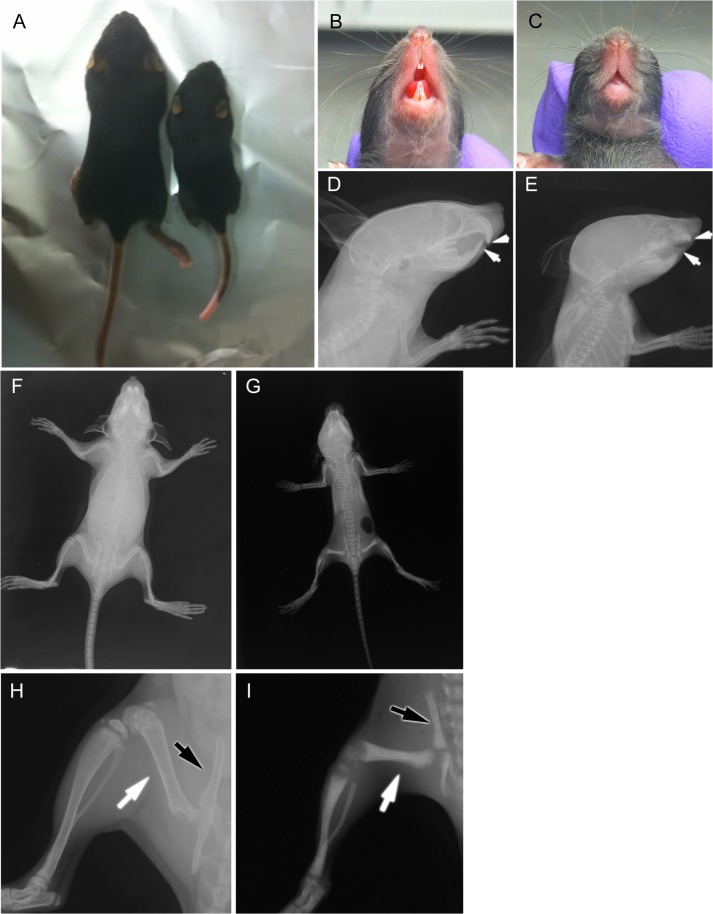
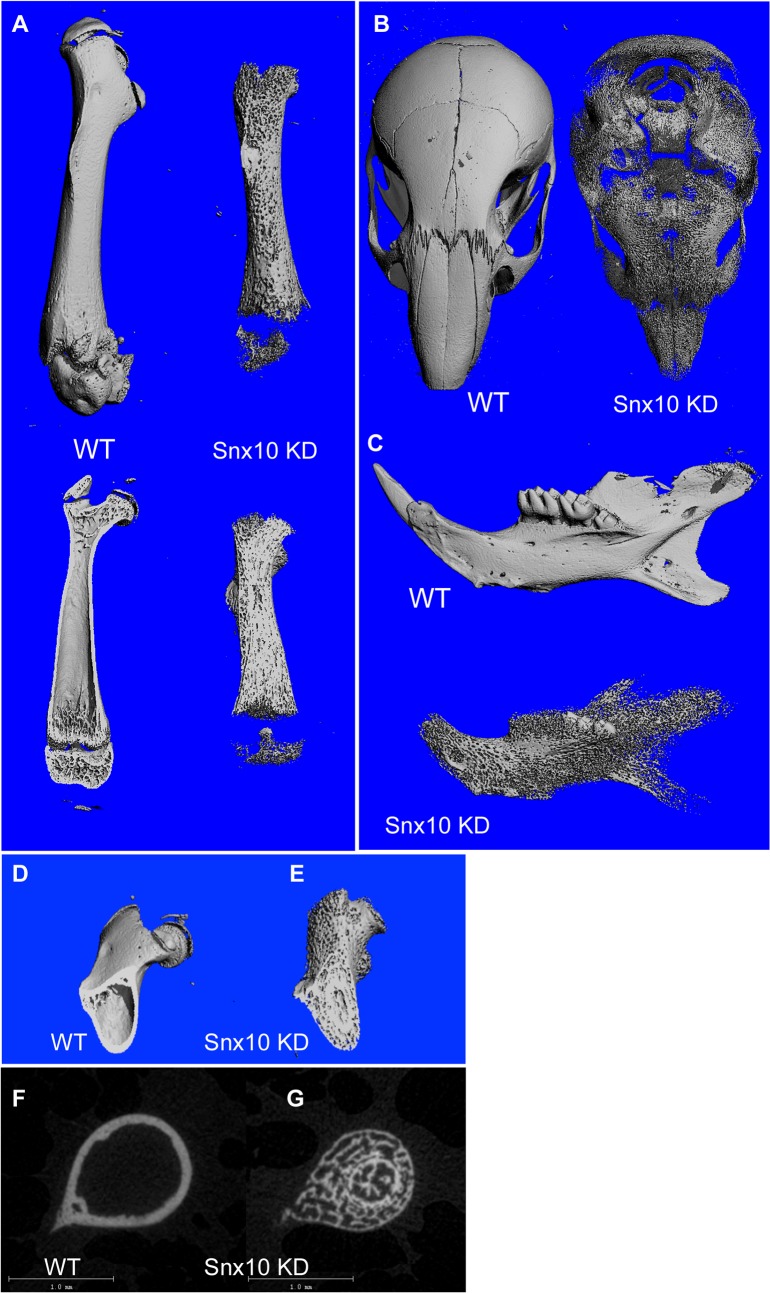
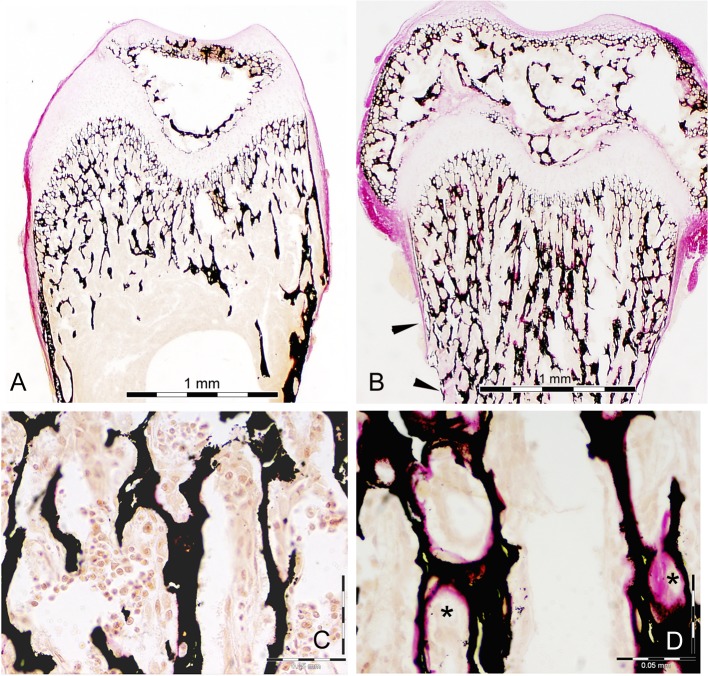
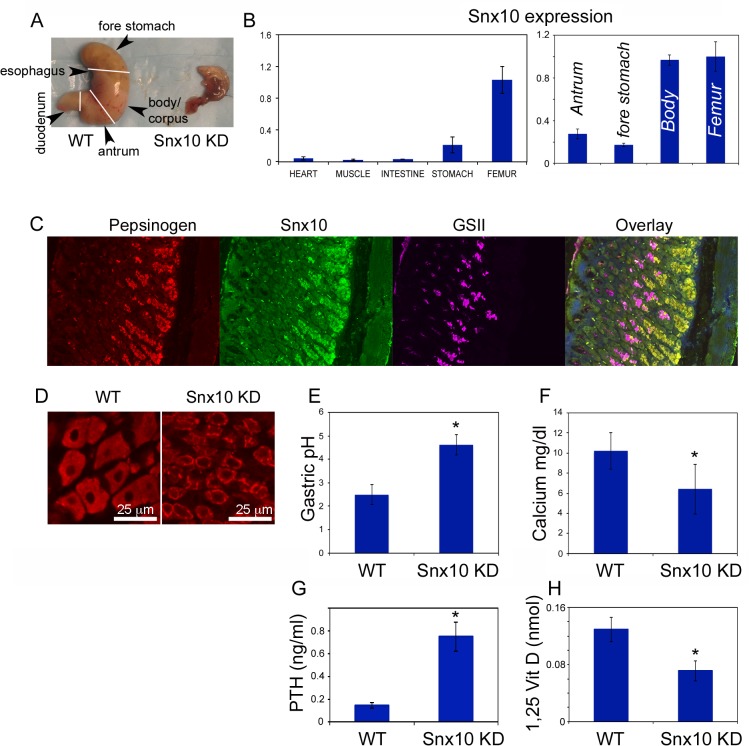
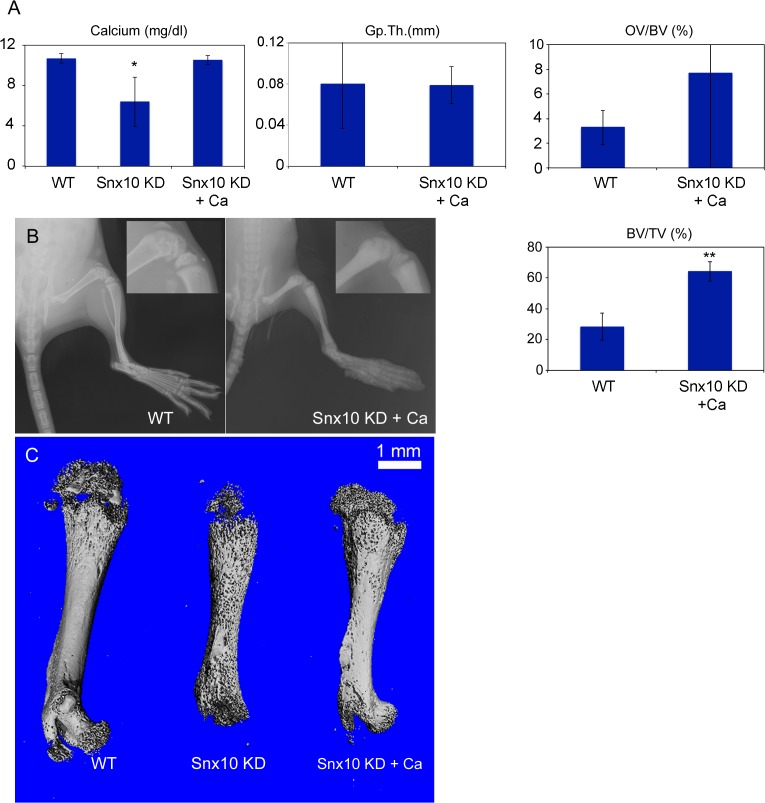
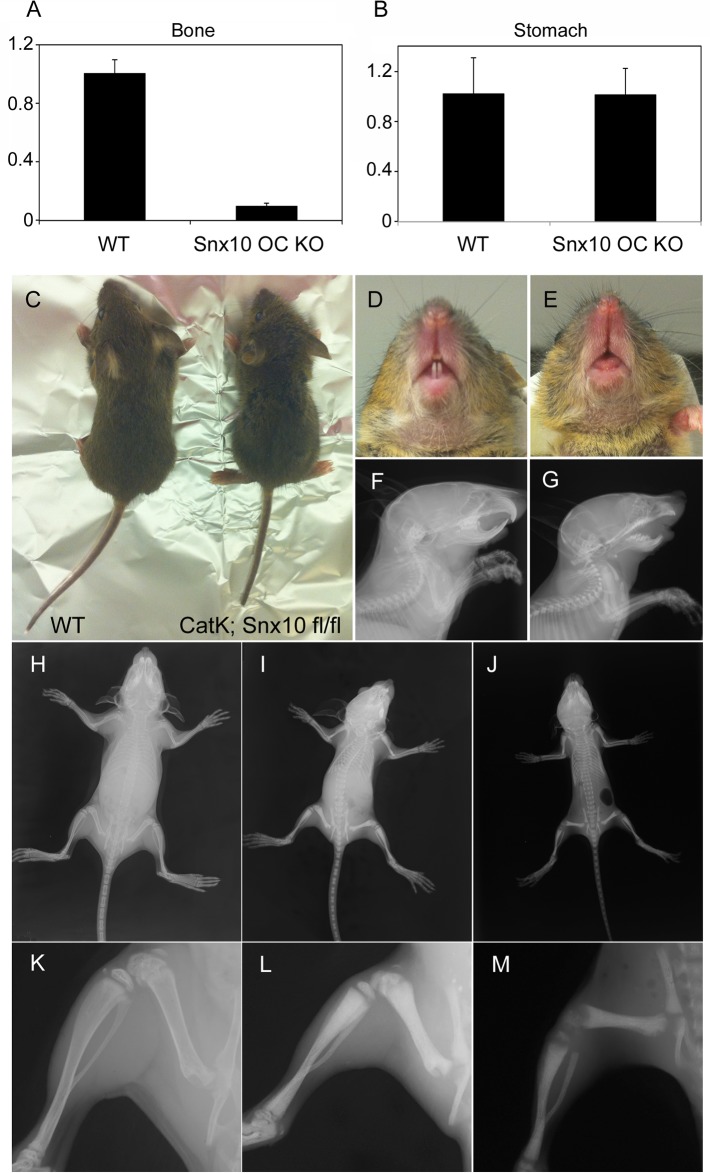
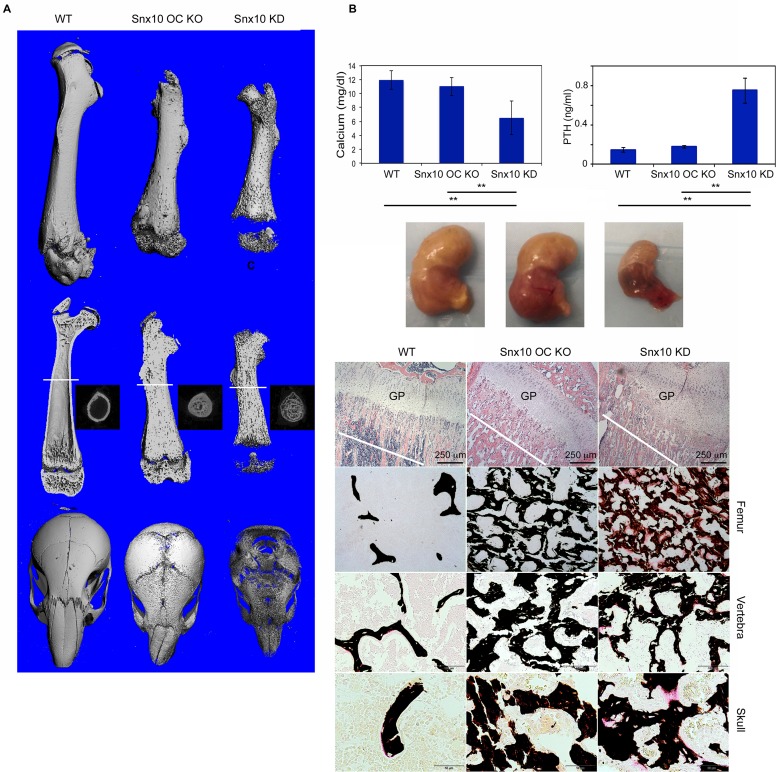
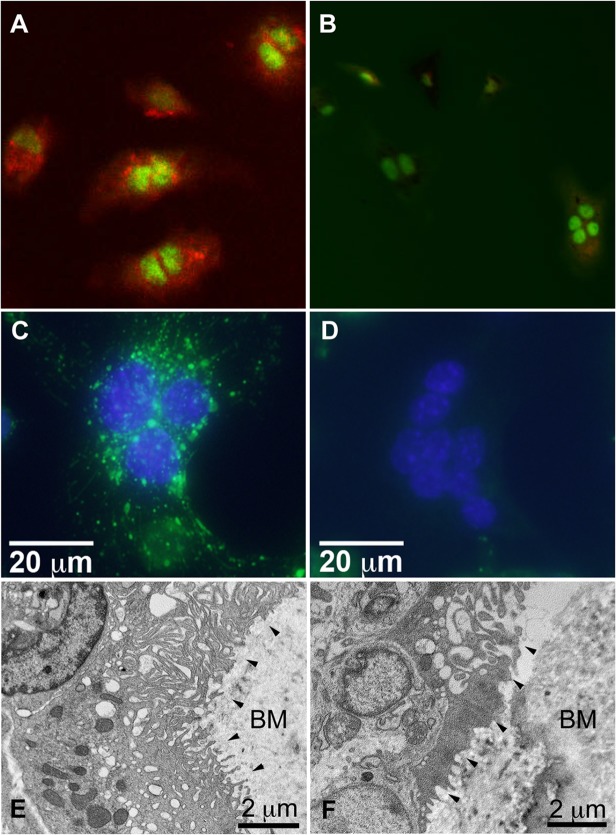
Mutations in sorting nexin 10 (Snx10) have recently been found to account for roughly 4% of all human malignant osteopetrosis, some of them fatal. To study the disease pathogenesis, we investigated the expression of Snx10 and created mouse models in which Snx10 was knocked down globally or knocked out in osteoclasts. Endocytosis is severely defective in Snx10-deficient osteoclasts, as is extracellular acidification, ruffled border formation, and bone resorption. We also discovered that Snx10 is highly expressed in stomach epithelium, with mutations leading to high stomach pH and low calcium solubilization. Global Snx10-deficiency in mice results in a combined phenotype: osteopetrosis (due to osteoclast defect) and rickets (due to high stomach pH and low calcium availability, resulting in impaired bone mineralization). Osteopetrorickets, the paradoxical association of insufficient mineralization in the context of a positive total body calcium balance, is thought to occur due to the inability of the osteoclasts to maintain normal calcium-phosphorus homeostasis. However, osteoclast-specific Snx10 knockout had no effect on calcium balance, and therefore led to severe osteopetrosis without rickets. Moreover, supplementation with calcium gluconate rescued mice from the rachitic phenotype and dramatically extended life span in global Snx10-deficient mice, suggesting that this may be a life-saving component of the clinical approach to Snx10-dependent human osteopetrosis that has previously gone unrecognized. We conclude that tissue-specific effects of Snx10 mutation need to be considered in clinical approaches to this disease entity. Reliance solely on hematopoietic stem cell transplantation can leave hypocalcemia uncorrected with sometimes fatal consequences. These studies established an essential role for Snx10 in bone homeostasis and underscore the importance of gastric acidification in calcium uptake.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
An SNX10 mutation causes malignant osteopetrosis of infancy.J Med Genet. 2012 Apr;49(4):221-6. doi: 10.1136/jmedgenet-2011-100520. J Med Genet. 2012. PMID: 22499339
-
SNX10 regulates osteoclastogenic cell fusion and osteoclast size in mice.J Bone Miner Res. 2024 Sep 26;39(10):1503-1517. doi: 10.1093/jbmr/zjae125. J Bone Miner Res. 2024. PMID: 39095084
-
Osteopetrosis associated with PLEKHM1 and SNX10 genes, both involved in osteoclast vesicular trafficking.Bone. 2022 Nov;164:116520. doi: 10.1016/j.bone.2022.116520. Epub 2022 Aug 15. Bone. 2022. PMID: 35981699
-
The molecular structure and function of sorting nexin 10 in skeletal disorders, cancers, and other pathological conditions.J Cell Physiol. 2021 Jun;236(6):4207-4215. doi: 10.1002/jcp.30173. Epub 2020 Nov 25. J Cell Physiol. 2021. PMID: 33241559 Review.
-
SNX10 gene mutation in infantile malignant osteopetrosis: A case report and literature review.Zhong Nan Da Xue Xue Bao Yi Xue Ban. 2021 Jan 28;46(1):108-112. doi: 10.11817/j.issn.1672-7347.2021.190322. Zhong Nan Da Xue Xue Bao Yi Xue Ban. 2021. PMID: 33678645 Free PMC article. Review. Chinese, English.
Cited by
-
Comparative transcriptome analysis of the main beam and brow tine of sika deer antler provides insights into the molecular control of rapid antler growth.Cell Mol Biol Lett. 2020 Sep 7;25:42. doi: 10.1186/s11658-020-00234-9. eCollection 2020. Cell Mol Biol Lett. 2020. PMID: 32944020 Free PMC article.
-
JAK2-IGF1 axis in osteoclasts regulates postnatal growth in mice.JCI Insight. 2021 Mar 8;6(5):e137045. doi: 10.1172/jci.insight.137045. JCI Insight. 2021. PMID: 33682794 Free PMC article.
-
Large-scale detection and characterization of interchromosomal rearrangements in normozoospermic bulls using massive genotype and phenotype data sets.Genome Res. 2023 Jun;33(6):957-971. doi: 10.1101/gr.277787.123. Epub 2023 Jul 6. Genome Res. 2023. PMID: 37414574 Free PMC article.
-
SNX10 gene mutation leading to osteopetrosis with dysfunctional osteoclasts.Sci Rep. 2017 Jun 7;7(1):3012. doi: 10.1038/s41598-017-02533-2. Sci Rep. 2017. PMID: 28592808 Free PMC article.
-
Absorption Characteristics of Novel Compound Calcium Carbonate Granules: Effects of Gastric Acid Deficiency and Exogenous Weak Acids.Curr Med Sci. 2019 Apr;39(2):337-342. doi: 10.1007/s11596-019-2040-4. Epub 2019 Apr 23. Curr Med Sci. 2019. PMID: 31016531
References
-
- Nesbitt SA, Horton MA (1997) Trafficking of matrix collagens through bone-resorbing osteoclasts. Science 276: 266–269. - PubMed
-
- Stenbeck G, Horton MA (2004) Endocytic trafficking in actively resorbing osteoclasts. J Cell Sci 117: 827–836. - PubMed
-
- Worby CA, Dixon JE (2002) Sorting out the cellular functions of sorting nexins. Nat Rev Mol Cell Biol 3: 919–931. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
