

Random mutagenesis identifies a C-terminal region of YopD important for Yersinia type III secretion function
- PMID: 25807250
- PMCID: PMC4433470
- DOI: 10.1371/journal.pone.0120471
Random mutagenesis identifies a C-terminal region of YopD important for Yersinia type III secretion function
Erratum in
-
Correction: Random Mutagenesis Identifies a C-Terminal Region of YopD Important for Yersinia Type III Secretion Function.PLoS One. 2015 Jun 12;10(6):e0130112. doi: 10.1371/journal.pone.0130112. eCollection 2015. PLoS One. 2015. PMID: 26068664 Free PMC article. No abstract available.
Abstract
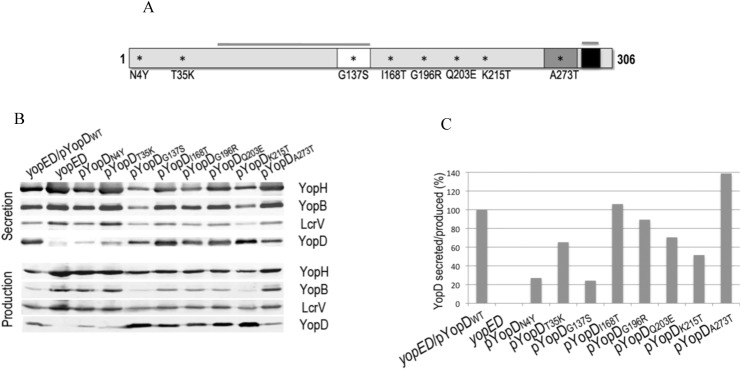
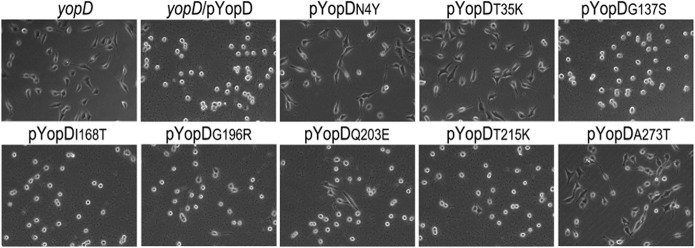
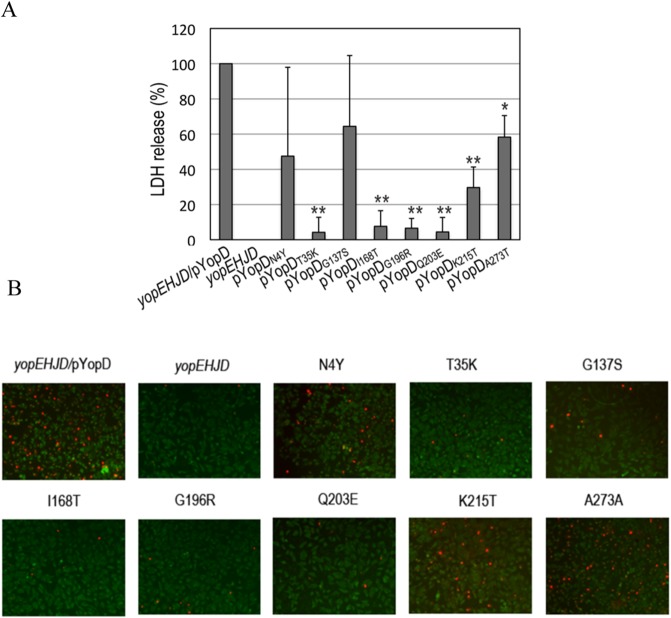
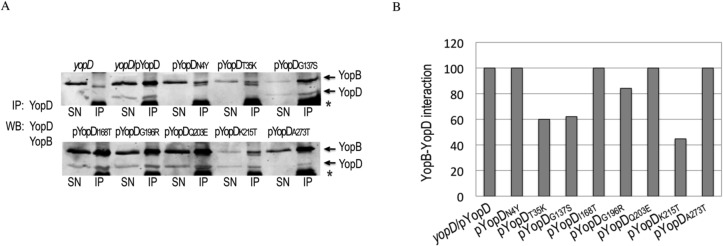
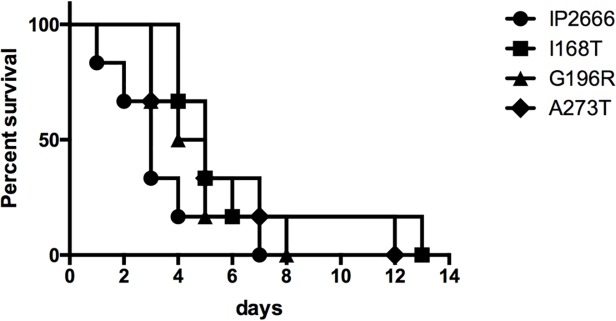
A common virulence mechanism among bacterial pathogens is the use of specialized secretion systems that deliver virulence proteins through a translocation channel inserted in the host cell membrane. During Yersinia infection, the host recognizes the type III secretion system mounting a pro-inflammatory response. However, soon after they are translocated, the effectors efficiently counteract that response. In this study we sought to identify YopD residues responsible for type III secretion system function. Through random mutagenesis, we identified eight Y. pseudotuberculosis yopD mutants with single amino acid changes affecting various type III secretion functions. Three severely defective mutants had substitutions in residues encompassing a 35 amino acid region (residues 168-203) located between the transmembrane domain and the C-terminal putative coiled-coil region of YopD. These mutations did not affect regulation of the low calcium response or YopB-YopD interaction but markedly inhibited MAPK and NFκB. [corrected] activation. When some of these mutations were introduced into the native yopD gene, defects in effector translocation and pore formation were also observed. We conclude that this newly identified region is important for YopD translocon function. The role of this domain in vivo remains elusive, as amino acid substitutions in that region did not significantly affect virulence of Y. pseudotuberculosis in orogastrically-infected mice.
Conflict of interest statement
Figures
Similar articles
-
YopD of Yersinia pseudotuberculosis is translocated into the cytosol of HeLa epithelial cells: evidence of a structural domain necessary for translocation.Mol Microbiol. 1998 Aug;29(3):799-813. doi: 10.1046/j.1365-2958.1998.00973.x. Mol Microbiol. 1998. PMID: 9723919
-
Yersinia pseudotuberculosis YopD mutants that genetically separate effector protein translocation from host membrane disruption.Mol Microbiol. 2015 May;96(4):764-78. doi: 10.1111/mmi.12970. Epub 2015 Mar 16. Mol Microbiol. 2015. PMID: 25684661
-
Coiled-coils in the YopD translocator family: a predicted structure unique to the YopD N-terminus contributes to full virulence of Yersinia pseudotuberculosis.Infect Genet Evol. 2012 Dec;12(8):1729-42. doi: 10.1016/j.meegid.2012.07.016. Epub 2012 Aug 11. Infect Genet Evol. 2012. PMID: 22910185
-
Yersinia outer proteins: role in modulation of host cell signaling responses and pathogenesis.Annu Rev Microbiol. 2005;59:69-89. doi: 10.1146/annurev.micro.59.030804.121320. Annu Rev Microbiol. 2005. PMID: 15847602 Review.
-
The Yersinia Yop virulon: a bacterial system for subverting eukaryotic cells.Mol Microbiol. 1997 Mar;23(5):861-7. doi: 10.1046/j.1365-2958.1997.2731623.x. Mol Microbiol. 1997. PMID: 9076724 Review.
Cited by
-
Yersinia versus host immunity: how a pathogen evades or triggers a protective response.Curr Opin Microbiol. 2016 Feb;29:56-62. doi: 10.1016/j.mib.2015.11.001. Epub 2015 Nov 27. Curr Opin Microbiol. 2016. PMID: 26638030 Free PMC article. Review.
-
Correction: Random Mutagenesis Identifies a C-Terminal Region of YopD Important for Yersinia Type III Secretion Function.PLoS One. 2015 Jun 12;10(6):e0130112. doi: 10.1371/journal.pone.0130112. eCollection 2015. PLoS One. 2015. PMID: 26068664 Free PMC article. No abstract available.
-
Macrophage innate immune responses delineate between defective translocon assemblies produced by Yersinia pseudotuberculosis YopD mutants.Virulence. 2023 Dec;14(1):2249790. doi: 10.1080/21505594.2023.2249790. Virulence. 2023. PMID: 37621095 Free PMC article.
-
Bacterial internalization is required to trigger NIK-dependent NF-κB activation in response to the bacterial type three secretion system.PLoS One. 2017 Feb 6;12(2):e0171406. doi: 10.1371/journal.pone.0171406. eCollection 2017. PLoS One. 2017. PMID: 28166267 Free PMC article.
References
-
- Viboud GI, Bliska JB. Yersinia outer proteins: role in modulation of host cell signaling responses and pathogenesis. Annu Rev Microbiol. 2005;59:69–89. - PubMed
-
- Sory MP, Cornelis GR. Translocation of a hybrid YopE-adenylate cyclase from Yersinia enterocolitica into HeLa cells. Mol Microbiol. 1994. Nov;14(3):583–94. - PubMed
-
- Persson C, Nordfelth R, Holmstrom A, Hakansson S, Rosqvist R, Wolf-Watz H. Cell-surface-bound Yersinia translocate the protein tyrosine phosphatase YopH by a polarized mechanism into the target cell. Mol Microbiol. 1995. Oct;18(1):135–50. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
