

SLAM- and nectin-4-independent noncytolytic spread of canine distemper virus in astrocytes
- PMID: 25787275
- PMCID: PMC4442543
- DOI: 10.1128/JVI.00004-15
SLAM- and nectin-4-independent noncytolytic spread of canine distemper virus in astrocytes
Abstract
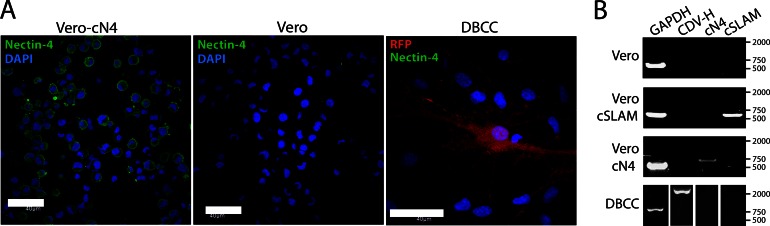
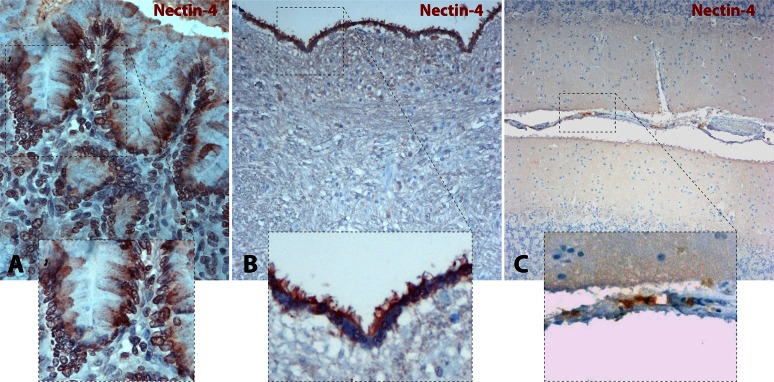
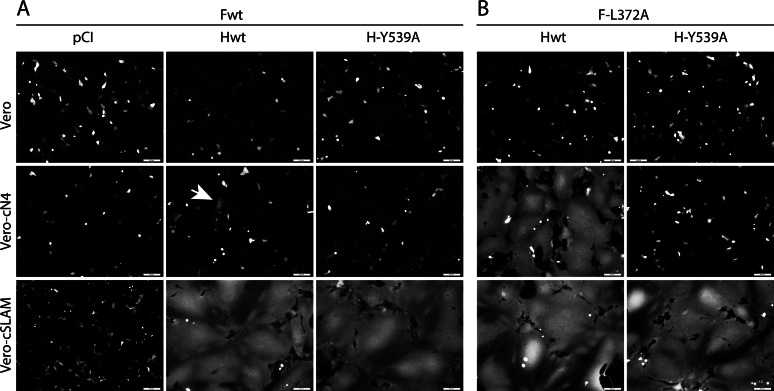
Measles and canine distemper viruses (MeV and CDV, respectively) first replicate in lymphatic and epithelial tissues by using SLAM and nectin-4 as entry receptors, respectively. The viruses may also invade the brain to establish persistent infections, triggering fatal complications, such as subacute sclerosis pan-encephalitis (SSPE) in MeV infection or chronic, multiple sclerosis-like, multifocal demyelinating lesions in the case of CDV infection. In both diseases, persistence is mediated by viral nucleocapsids that do not require packaging into particles for infectivity but are directly transmitted from cell to cell (neurons in SSPE or astrocytes in distemper encephalitis), presumably by relying on restricted microfusion events. Indeed, although morphological evidence of fusion remained undetectable, viral fusion machineries and, thus, a putative cellular receptor, were shown to contribute to persistent infections. Here, we first showed that nectin-4-dependent cell-cell fusion in Vero cells, triggered by a demyelinating CDV strain, remained extremely limited, thereby supporting a potential role of nectin-4 in mediating persistent infections in astrocytes. However, nectin-4 could not be detected in either primary cultured astrocytes or the white matter of tissue sections. In addition, a bioengineered "nectin-4-blind" recombinant CDV retained full cell-to-cell transmission efficacy in primary astrocytes. Combined with our previous report demonstrating the absence of SLAM expression in astrocytes, these findings are suggestive for the existence of a hitherto unrecognized third CDV receptor expressed by glial cells that contributes to the induction of noncytolytic cell-to-cell viral transmission in astrocytes.
Importance: While persistent measles virus (MeV) infection induces SSPE in humans, persistent canine distemper virus (CDV) infection causes chronic progressive or relapsing demyelination in carnivores. Common to both central nervous system (CNS) infections is that persistence is based on noncytolytic cell-to-cell spread, which, in the case of CDV, was demonstrated to rely on functional membrane fusion machinery complexes. This inferred a mechanism where nucleocapsids are transmitted through macroscopically invisible microfusion events between infected and target cells. Here, we provide evidence that CDV induces such microfusions in a SLAM- and nectin-4-independent manner, thereby strongly suggesting the existence of a third receptor expressed in glial cells (referred to as GliaR). We propose that GliaR governs intercellular transfer of nucleocapsids and hence contributes to viral persistence in the brain and ensuing demyelinating lesions.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
Comment in
-
Commentary: SLAM- and Nectin-4-Independent Noncytolytic Spread of Canine Distemper Virus in Astrocytes.Front Microbiol. 2016 Dec 9;7:2011. doi: 10.3389/fmicb.2016.02011. eCollection 2016. Front Microbiol. 2016. PMID: 28018332 Free PMC article. No abstract available.
Similar articles
-
Infectious Progression of Canine Distemper Virus from Circulating Cerebrospinal Fluid into the Central Nervous System.J Virol. 2016 Sep 29;90(20):9285-92. doi: 10.1128/JVI.01337-16. Print 2016 Oct 15. J Virol. 2016. PMID: 27489268 Free PMC article.
-
Canine Distemper Virus Spread and Transmission to Naive Ferrets: Selective Pressure on Signaling Lymphocyte Activation Molecule-Dependent Entry.J Virol. 2018 Jul 17;92(15):e00669-18. doi: 10.1128/JVI.00669-18. Print 2018 Aug 1. J Virol. 2018. PMID: 29793948 Free PMC article.
-
Experimental adaptation of wild-type canine distemper virus (CDV) to the human entry receptor CD150.PLoS One. 2013;8(3):e57488. doi: 10.1371/journal.pone.0057488. Epub 2013 Mar 12. PLoS One. 2013. PMID: 23554862 Free PMC article.
-
Multiple Receptors Involved in Invasion and Neuropathogenicity of Canine Distemper Virus: A Review.Viruses. 2022 Jul 12;14(7):1520. doi: 10.3390/v14071520. Viruses. 2022. PMID: 35891500 Free PMC article. Review.
-
Tropism and molecular pathogenesis of canine distemper virus.Virol J. 2019 Mar 7;16(1):30. doi: 10.1186/s12985-019-1136-6. Virol J. 2019. PMID: 30845967 Free PMC article. Review.
Cited by
-
Host Cellular Receptors for the Peste des Petits Ruminant Virus.Viruses. 2019 Aug 8;11(8):729. doi: 10.3390/v11080729. Viruses. 2019. PMID: 31398809 Free PMC article. Review.
-
Cetacean Host-Pathogen Interaction(s): Critical Knowledge Gaps.Front Immunol. 2018 Nov 28;9:2815. doi: 10.3389/fimmu.2018.02815. eCollection 2018. Front Immunol. 2018. PMID: 30546370 Free PMC article. Review. No abstract available.
-
Evolution and Interspecies Transmission of Canine Distemper Virus-An Outlook of the Diverse Evolutionary Landscapes of a Multi-Host Virus.Viruses. 2019 Jun 26;11(7):582. doi: 10.3390/v11070582. Viruses. 2019. PMID: 31247987 Free PMC article. Review.
-
Infectious Progression of Canine Distemper Virus from Circulating Cerebrospinal Fluid into the Central Nervous System.J Virol. 2016 Sep 29;90(20):9285-92. doi: 10.1128/JVI.01337-16. Print 2016 Oct 15. J Virol. 2016. PMID: 27489268 Free PMC article.
-
First report of canine morbillivirus infection of adipose tissue-derived stem cells from dogs with distemper.Vet World. 2022 Jul;15(7):1835-1842. doi: 10.14202/vetworld.2022.1835-1842. Epub 2022 Jul 27. Vet World. 2022. PMID: 36185532 Free PMC article.
References
-
- Leonard VH, Hodge G, Reyes-Del VJ, McChesney MB, Cattaneo R. 2010. Measles virus selectively blind to signaling lymphocytic activation molecule (SLAM; CD150) is attenuated and induces strong adaptive immune responses in rhesus monkeys. J Virol 84:3413–3420. doi:10.1128/JVI.02304-09. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
