

Light-sensitive Phytochrome-Interacting Factors (PIFs) are not required to regulate phytoene synthase gene expression in the root
- PMID: 25763615
- PMCID: PMC4203534
- DOI: 10.4161/psb.29248
Light-sensitive Phytochrome-Interacting Factors (PIFs) are not required to regulate phytoene synthase gene expression in the root
Abstract
Carotenoids are plastidial isoprenoids essential for the protection of photosynthetic tissues against excess light. They also serve as precursors of apocarotenoid hormones such as abscisic acid (ABA) and strigolactones. The first enzyme of the carotenoid pathway, phytoene synthase (PSY), is also the main rate-limiting step. Unlike that observed in most plants, PSY is encoded by a single gene in Arabidopsis thaliana. Whereas the PSY gene is induced by light in photosynthetic tissues, a root-specific upregulation of PSY expression by salt stress and ABA has been recently demonstrated. Here we report that transcription factors of the Phytochrome-Interacting Factor (PIF) family, previously shown to repress PSY expression in etiolated seedlings and mature leaves, do not influence PSY expression in roots. Together, our results suggest that organ-specific pathways regulate PSY expression and hence carotenoid production in response to different environmental cues.
Keywords: Arabidopsis; PIF; carotenoid; dark; phytoene synthase; root; salt.
Figures
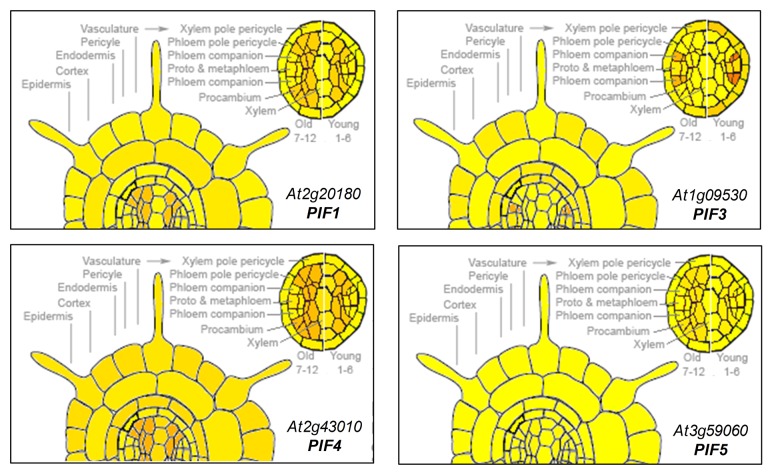
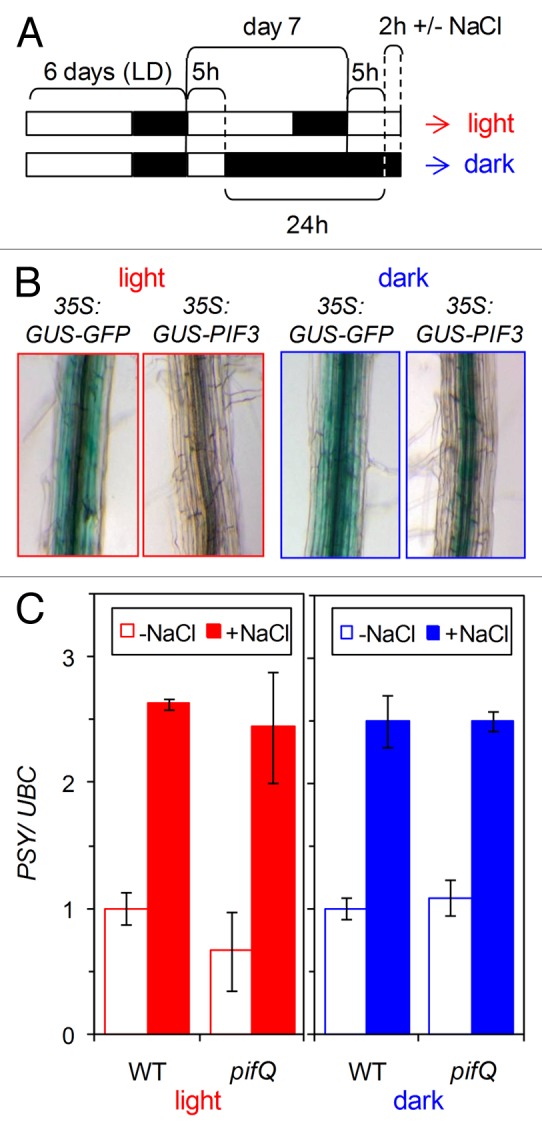
Similar articles
-
Direct regulation of phytoene synthase gene expression and carotenoid biosynthesis by phytochrome-interacting factors.Proc Natl Acad Sci U S A. 2010 Jun 22;107(25):11626-31. doi: 10.1073/pnas.0914428107. Epub 2010 Jun 7. Proc Natl Acad Sci U S A. 2010. PMID: 20534526 Free PMC article.
-
Light-dependent regulation of carotenoid biosynthesis occurs at the level of phytoene synthase expression and is mediated by phytochrome in Sinapis alba and Arabidopsis thaliana seedlings.Plant J. 1997 Sep;12(3):625-34. doi: 10.1046/j.1365-313x.1997.00625.x. Plant J. 1997. PMID: 9351247
-
PSY3, a new member of the phytoene synthase gene family conserved in the Poaceae and regulator of abiotic stress-induced root carotenogenesis.Plant Physiol. 2008 Mar;146(3):1333-45. doi: 10.1104/pp.107.111120. Epub 2007 Dec 27. Plant Physiol. 2008. PMID: 18162592 Free PMC article.
-
Evolution of root-specific carotenoid precursor pathways for apocarotenoid signal biogenesis.Plant Sci. 2015 Apr;233:1-10. doi: 10.1016/j.plantsci.2014.12.017. Epub 2014 Dec 30. Plant Sci. 2015. PMID: 25711808 Review.
-
Functions of Phytochrome-Interacting Factors (PIFs) in the regulation of plant growth and development: A comprehensive review.Int J Biol Macromol. 2023 Jul 31;244:125234. doi: 10.1016/j.ijbiomac.2023.125234. Epub 2023 Jun 7. Int J Biol Macromol. 2023. PMID: 37290549 Review.
Cited by
-
Chloroplast-to-chromoplast transition envisions provitamin A biofortification in green vegetables.Plant Cell Rep. 2021 May;40(5):799-804. doi: 10.1007/s00299-021-02684-7. Epub 2021 Mar 23. Plant Cell Rep. 2021. PMID: 33754204
-
Putative Daucus carota Capsanthin-Capsorubin Synthase (DcCCS) Possesses Lycopene β-Cyclase Activity, Boosts Carotenoid Levels, and Increases Salt Tolerance in Heterologous Plants.Plants (Basel). 2023 Jul 27;12(15):2788. doi: 10.3390/plants12152788. Plants (Basel). 2023. PMID: 37570943 Free PMC article.
-
Regulation of Carotenoid Biosynthesis by Shade Relies on Specific Subsets of Antagonistic Transcription Factors and Cofactors.Plant Physiol. 2015 Nov;169(3):1584-94. doi: 10.1104/pp.15.00552. Epub 2015 Jun 16. Plant Physiol. 2015. PMID: 26082398 Free PMC article.
-
Overexpression of PSY1 increases fruit skin and flesh carotenoid content and reveals associated transcription factors in apple (Malus × domestica).Front Plant Sci. 2022 Sep 15;13:967143. doi: 10.3389/fpls.2022.967143. eCollection 2022. Front Plant Sci. 2022. PMID: 36186009 Free PMC article.
References
-
- Botella-Pavía P, Rodríguez-Concepción M. Carotenoid biotechnology in plants for nutritionally improved foods. Physiol Plant. 2006;126:369–81. doi: 10.1111/j.1399-3054.2006.00632.x. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases