

The RED domain of Paired is specifically required for Drosophila accessory gland maturation
- PMID: 25694546
- PMCID: PMC4345280
- DOI: 10.1098/rsob.140179
The RED domain of Paired is specifically required for Drosophila accessory gland maturation
Abstract
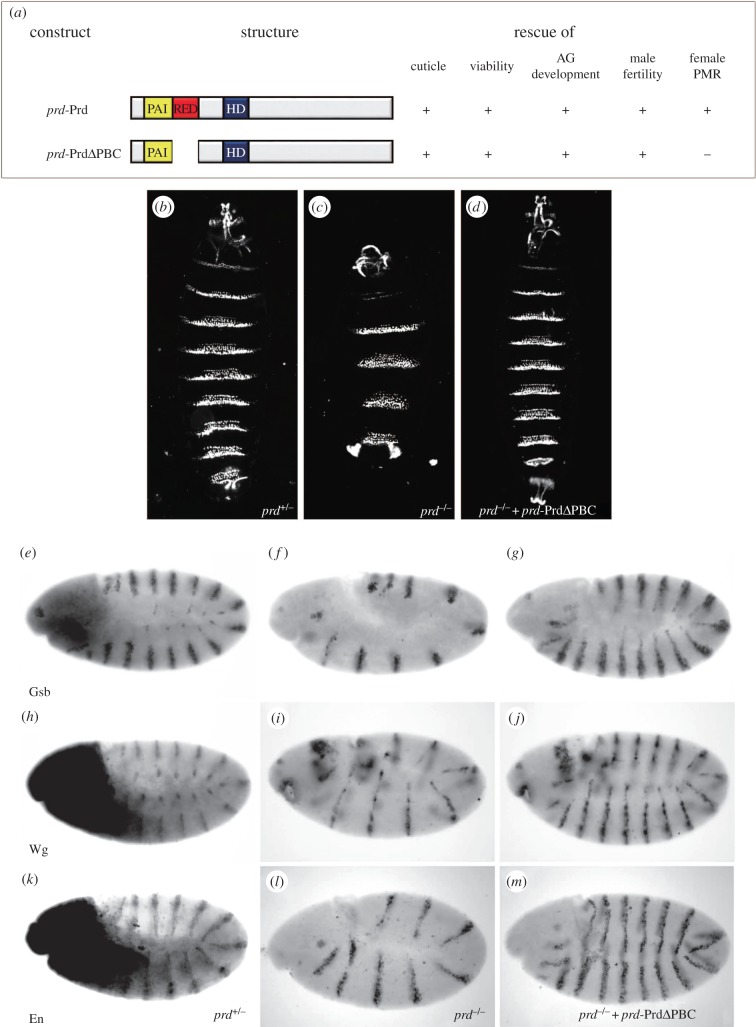
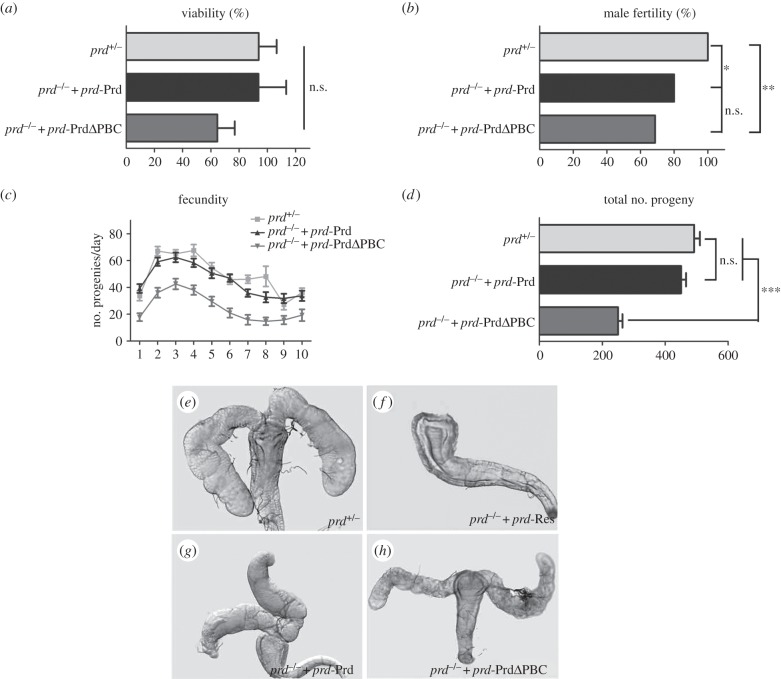
The evolutionarily conserved paired domain consists of the N-terminal PAI and the C-terminal RED domains, each containing a helix-turn-helix motif capable of binding DNA. Despite its conserved sequence, the physiological functions of the RED domain remain elusive. Here, we constructed a prd transgene expressing a truncated Paired (Prd) protein without the RED domain, and examined its rescue ability in prd mutants. We found that the RED domain is specifically required for the expression of Acp26Aa and sex peptide in male accessory glands, and the induction of female post-mating response. Our data thus identified an important physiological function for the evolutionarily conserved RED domain.
Keywords: Drosophila; Paired; RED domain; accessory gland; post-mating response.
Figures


Similar articles
-
In vivo requirement for the paired domain and homeodomain of the paired segmentation gene product.Development. 1996 Sep;122(9):2673-85. doi: 10.1242/dev.122.9.2673. Development. 1996. PMID: 8787742
-
Dual role of the Pax gene paired in accessory gland development of Drosophila.Development. 2002 Jan;129(2):339-46. doi: 10.1242/dev.129.2.339. Development. 2002. PMID: 11807027
-
A novel function for the Hox gene Abd-B in the male accessory gland regulates the long-term female post-mating response in Drosophila.PLoS Genet. 2013 Mar;9(3):e1003395. doi: 10.1371/journal.pgen.1003395. Epub 2013 Mar 28. PLoS Genet. 2013. PMID: 23555301 Free PMC article.
-
Functional male accessory glands and fertility in Drosophila require novel ecdysone receptor.PLoS Genet. 2017 May 11;13(5):e1006788. doi: 10.1371/journal.pgen.1006788. eCollection 2017 May. PLoS Genet. 2017. PMID: 28493870 Free PMC article.
-
Multiple protein functions of paired in Drosophila development and their conservation in the Gooseberry and Pax3 homologs.Development. 2001 Feb;128(3):395-405. doi: 10.1242/dev.128.3.395. Development. 2001. PMID: 11152638
Cited by
-
Analysis of the Anti-Tumour Effect of Xuefu Zhuyu Decoction Based on Network Pharmacology and Experimental Verification in Drosophila.Front Pharmacol. 2022 Jul 12;13:922457. doi: 10.3389/fphar.2022.922457. eCollection 2022. Front Pharmacol. 2022. PMID: 35903326 Free PMC article.
References
-
- Nusslein-Volhard C, Wieschaus E. 1980. Mutations affecting segment number and polarity in Drosophila. Nature 287, 795–801 (doi:10.1038/287795a0) - DOI - PubMed
-
- Baumgartner S, Noll M. 1990. Network of interactions among pair-rule genes regulating paired expression during primordial segmentation of Drosophila. Mech. Dev. 33, 1–18 (doi:10.1016/0925-4773(90)90130-E) - DOI - PubMed
-
- Ingham PW, Martinez Arias A. 1992. Boundaries and fields in early embryos. Cell 68, 221–235 (doi:10.1016/0092-8674(92)90467-Q) - DOI - PubMed
-
- St Johnston D, Nusslein-Volhard C. 1992. The origin of pattern and polarity in the Drosophila embryo. Cell 68, 201–219 (doi:10.1016/0092-8674(92)90466-P) - DOI - PubMed
-
- DiNardo S, O'Farrell PH. 1987. Establishment and refinement of segmental pattern in the Drosophila embryo: spatial control of engrailed expression by pair-rule genes. Genes Dev. 1, 1212–1225 (doi:10.1101/gad.1.10.1212) - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
