

A nucleotide exchange factor promotes endoplasmic reticulum-to-cytosol membrane penetration of the nonenveloped virus simian virus 40
- PMID: 25653441
- PMCID: PMC4442351
- DOI: 10.1128/JVI.03552-14
A nucleotide exchange factor promotes endoplasmic reticulum-to-cytosol membrane penetration of the nonenveloped virus simian virus 40
Abstract
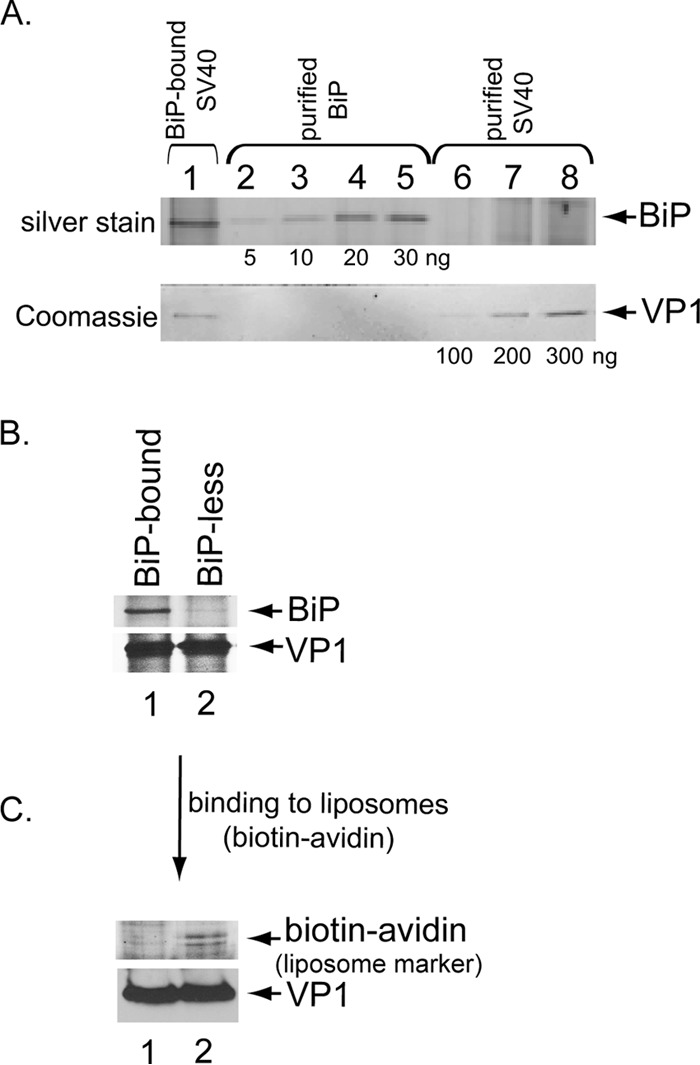
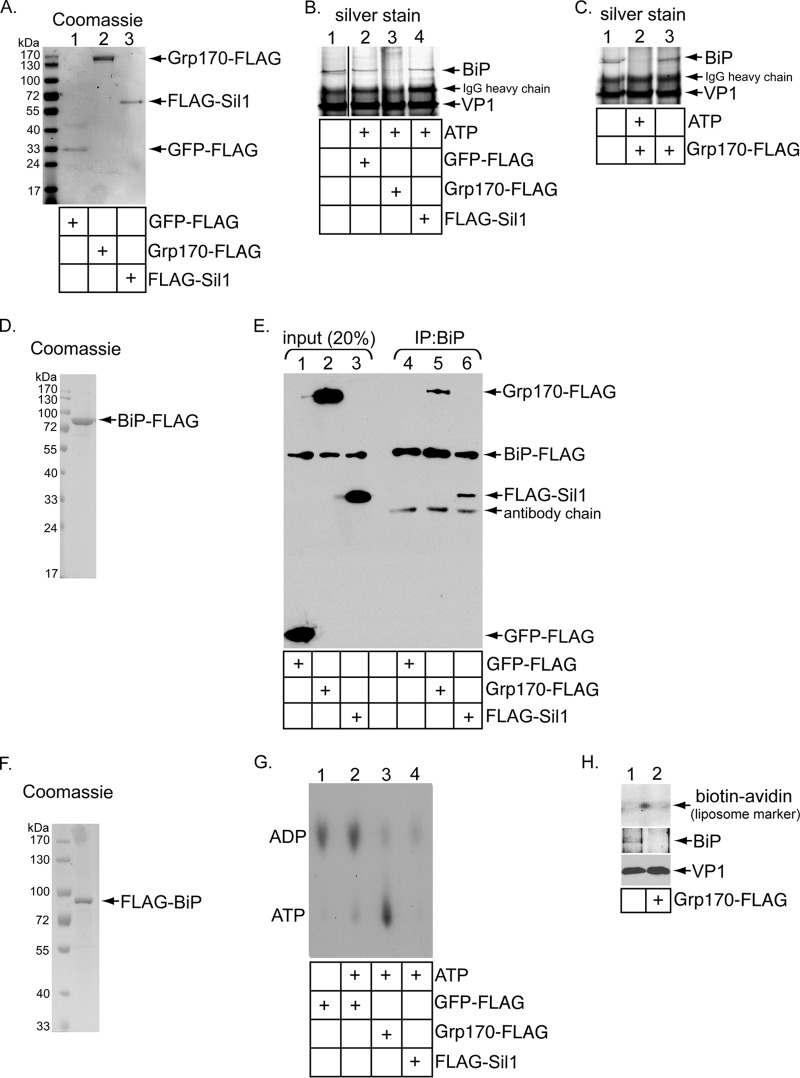
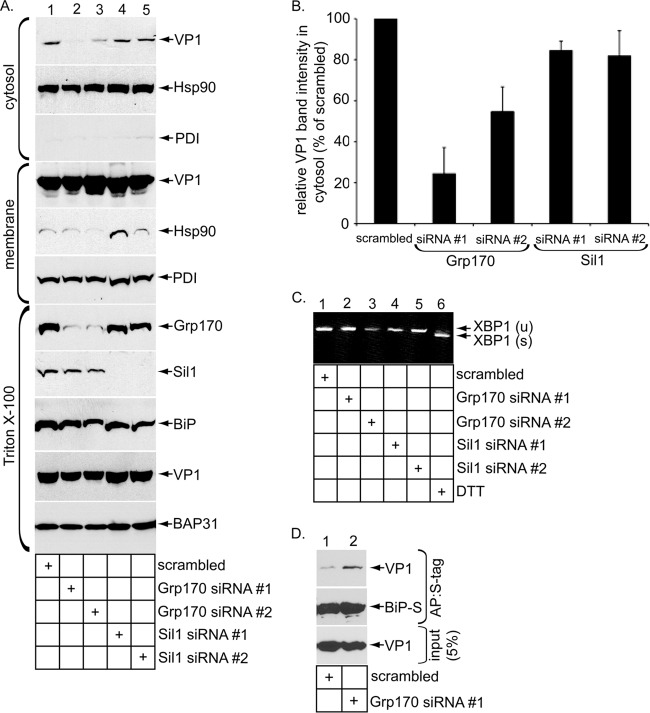
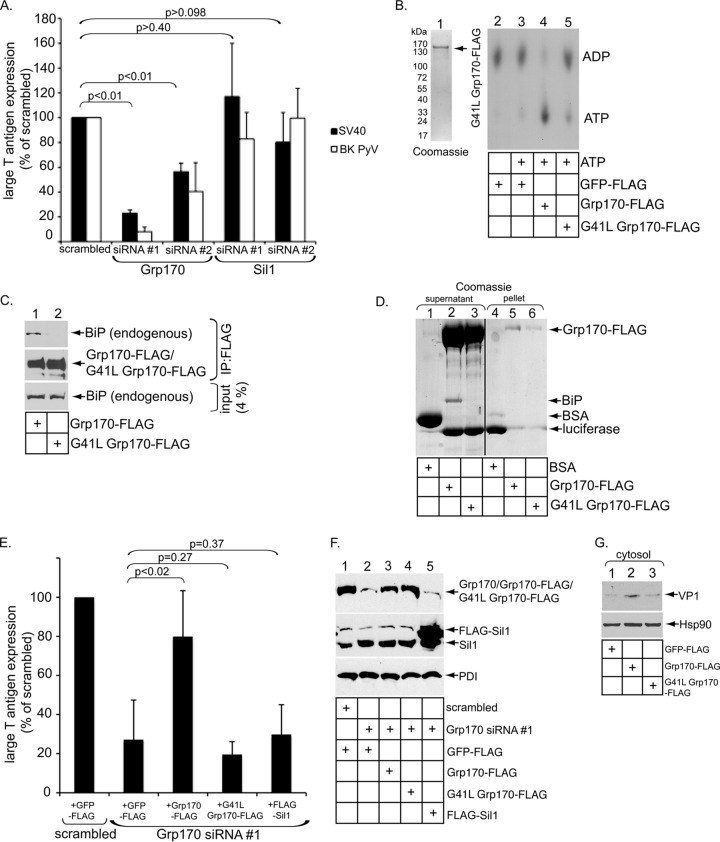
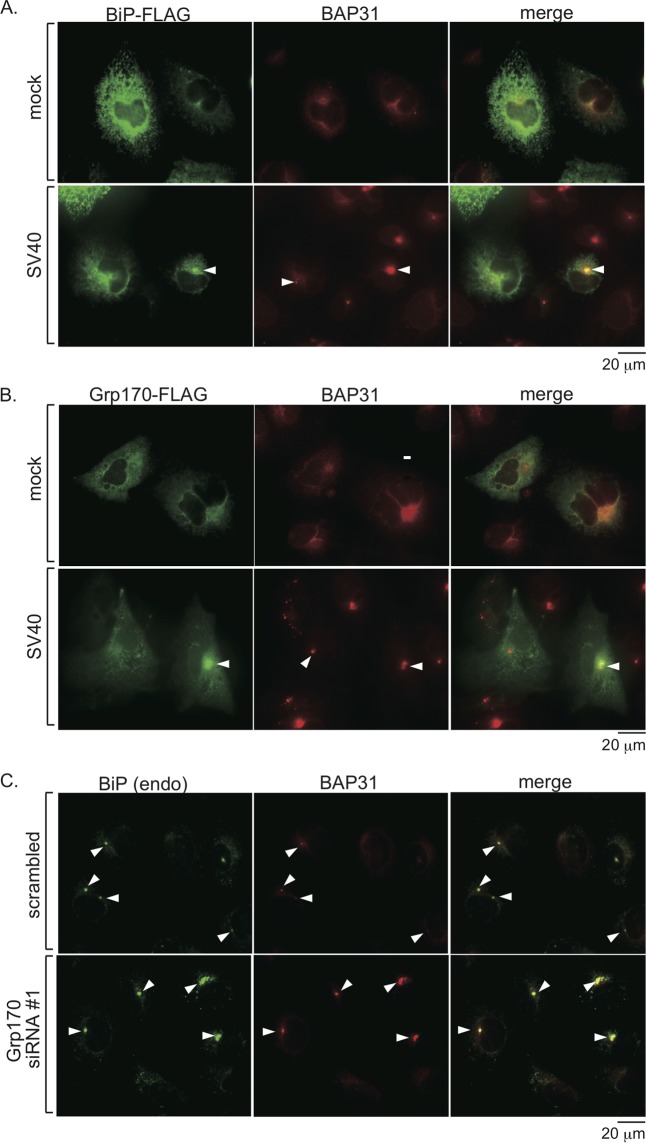
The nonenveloped simian polyomavirus (PyV) simian virus 40 (SV40) hijacks the endoplasmic reticulum (ER) quality control machinery to penetrate the ER membrane and reach the cytosol, a critical infection step. During entry, SV40 traffics to the ER, where host-induced conformational changes render the virus hydrophobic. The hydrophobic virus binds and integrates into the ER lipid bilayer to initiate membrane penetration. However, prior to membrane transport, the hydrophobic SV40 recruits the ER-resident Hsp70 BiP, which holds the virus in a transport-competent state until it is ready to cross the ER membrane. Here we probed how BiP disengages from SV40 to enable the virus to penetrate the ER membrane. We found that nucleotide exchange factor (NEF) Grp170 induces nucleotide exchange of BiP and releases SV40 from BiP. Importantly, this reaction promotes SV40 ER-to-cytosol transport and infection. The human BK PyV also relies on Grp170 for successful infection. Interestingly, SV40 mobilizes a pool of Grp170 into discrete puncta in the ER called foci. These foci, postulated to represent the ER membrane penetration site, harbor ER components, including BiP, known to facilitate viral ER-to-cytosol transport. Our results thus identify a nucleotide exchange activity essential for catalyzing the most proximal event before ER membrane penetration of PyVs.
Importance: PyVs are known to cause debilitating human diseases. During entry, this virus family, including monkey SV40 and human BK PyV, hijacks ER protein quality control machinery to breach the ER membrane and access the cytosol, a decisive infection step. In this study, we pinpointed an ER-resident factor that executes a crucial role in promoting ER-to-cytosol membrane penetration of PyVs. Identifying a host factor that facilitates entry of the PyV family thus provides additional therapeutic targets to combat PyV-induced diseases.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
Similar articles
-
The endoplasmic reticulum membrane J protein C18 executes a distinct role in promoting simian virus 40 membrane penetration.J Virol. 2015 Apr;89(8):4058-68. doi: 10.1128/JVI.03574-14. Epub 2015 Jan 28. J Virol. 2015. PMID: 25631089 Free PMC article.
-
Bag2 Is a Component of a Cytosolic Extraction Machinery That Promotes Membrane Penetration of a Nonenveloped Virus.J Virol. 2018 Jul 17;92(15):e00607-18. doi: 10.1128/JVI.00607-18. Print 2018 Aug 1. J Virol. 2018. PMID: 29769335 Free PMC article.
-
SGTA-Dependent Regulation of Hsc70 Promotes Cytosol Entry of Simian Virus 40 from the Endoplasmic Reticulum.J Virol. 2017 May 26;91(12):e00232-17. doi: 10.1128/JVI.00232-17. Print 2017 Jun 15. J Virol. 2017. PMID: 28356524 Free PMC article.
-
SV40 Hijacks Cellular Transport, Membrane Penetration, and Disassembly Machineries to Promote Infection.Viruses. 2019 Oct 5;11(10):917. doi: 10.3390/v11100917. Viruses. 2019. PMID: 31590347 Free PMC article. Review.
-
How Polyomaviruses Exploit the ERAD Machinery to Cause Infection.Viruses. 2016 Aug 29;8(9):242. doi: 10.3390/v8090242. Viruses. 2016. PMID: 27589785 Free PMC article. Review.
Cited by
-
BiP and its nucleotide exchange factors Grp170 and Sil1: mechanisms of action and biological functions.J Mol Biol. 2015 Apr 10;427(7):1589-608. doi: 10.1016/j.jmb.2015.02.011. Epub 2015 Feb 16. J Mol Biol. 2015. PMID: 25698114 Free PMC article. Review.
-
A bacterial toxin and a nonenveloped virus hijack ER-to-cytosol membrane translocation pathways to cause disease.Crit Rev Biochem Mol Biol. 2015;50(6):477-88. doi: 10.3109/10409238.2015.1085826. Epub 2015 Sep 11. Crit Rev Biochem Mol Biol. 2015. PMID: 26362261 Free PMC article. Review.
-
The nucleotide exchange factors Grp170 and Sil1 induce cholera toxin release from BiP to enable retrotranslocation.Mol Biol Cell. 2015 Jun 15;26(12):2181-9. doi: 10.1091/mbc.E15-01-0014. Epub 2015 Apr 15. Mol Biol Cell. 2015. PMID: 25877869 Free PMC article.
-
Proteostasis in Viral Infection: Unfolding the Complex Virus-Chaperone Interplay.Cold Spring Harb Perspect Biol. 2020 Mar 2;12(3):a034090. doi: 10.1101/cshperspect.a034090. Cold Spring Harb Perspect Biol. 2020. PMID: 30858229 Free PMC article. Review.
-
Identification of Rab18 as an Essential Host Factor for BK Polyomavirus Infection Using a Whole-Genome RNA Interference Screen.mSphere. 2017 Jul 26;2(4):e00291-17. doi: 10.1128/mSphereDirect.00291-17. eCollection 2017 Jul-Aug. mSphere. 2017. PMID: 28815213 Free PMC article.
References
-
- Goodwin EC, Lipovsky A, Inoue T, Magaldi TG, Edwards AP, Van Goor KE, Paton AW, Paton JC, Atwood WJ, Tsai B, DiMaio D. 2011. BiP and multiple DNAJ molecular chaperones in the endoplasmic reticulum are required for efficient simian virus 40 infection. mBio 2:e00101-11. doi:10.1128/mBio.00101-11. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
