

Lipid agonism: The PIP2 paradigm of ligand-gated ion channels
- PMID: 25633344
- PMCID: PMC4540326
- DOI: 10.1016/j.bbalip.2015.01.011
Lipid agonism: The PIP2 paradigm of ligand-gated ion channels
Abstract
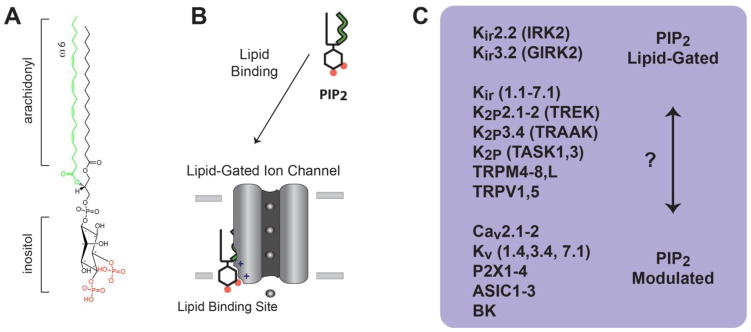
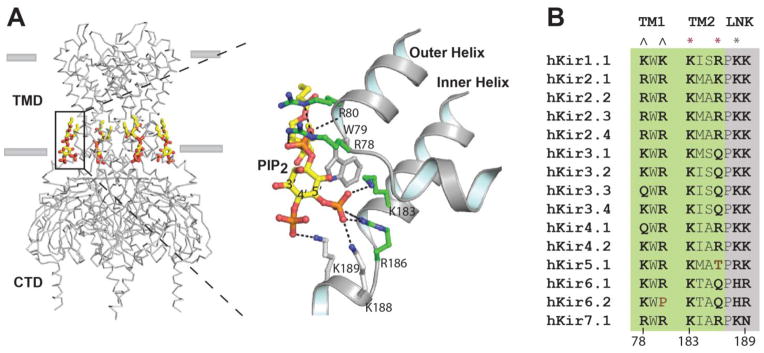
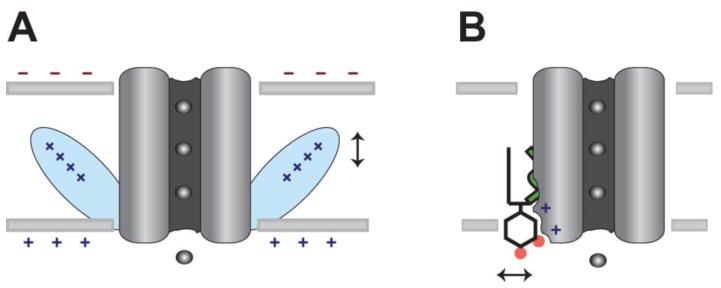
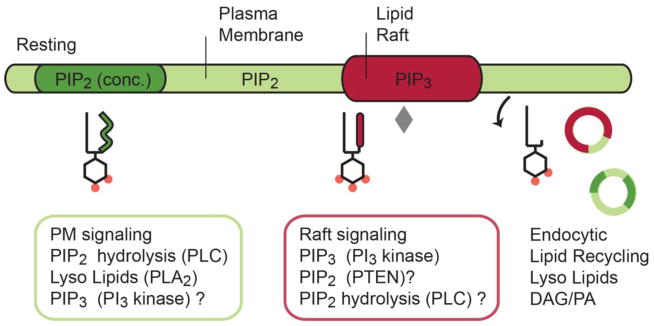
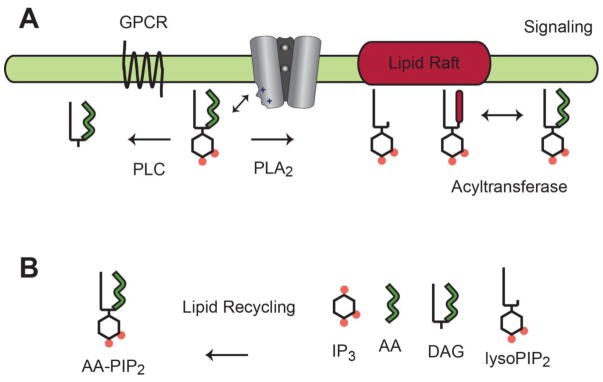
The past decade, membrane signaling lipids emerged as major regulators of ion channel function. However, the molecular nature of lipid binding to ion channels remained poorly described due to a lack of structural information and assays to quantify and measure lipid binding in a membrane. How does a lipid-ligand bind to a membrane protein in the plasma membrane, and what does it mean for a lipid to activate or regulate an ion channel? How does lipid binding compare to activation by soluble neurotransmitter? And how does the cell control lipid agonism? This review focuses on lipids and their interactions with membrane proteins, in particular, ion channels. I discuss the intersection of membrane lipid biology and ion channel biophysics. A picture emerges of membrane lipids as bona fide agonists of ligand-gated ion channels. These freely diffusing signals reside in the plasma membrane, bind to the transmembrane domain of protein, and cause a conformational change that allosterically gates an ion channel. The system employs a catalog of diverse signaling lipids ultimately controlled by lipid enzymes and raft localization. I draw upon pharmacology, recent protein structure, and electrophysiological data to understand lipid regulation and define inward rectifying potassium channels (Kir) as a new class of PIP2 lipid-gated ion channels.
Keywords: G-protein; Ion channel; Lipid gated; Lipid raft; PIP(2); Signaling lipid.
Copyright © 2015. Published by Elsevier B.V.
Conflict of interest statement
Conflict of interest.
The author declares no conflict of interest.
Figures
Similar articles
-
Structural basis of PIP2 activation of the classical inward rectifier K+ channel Kir2.2.Nature. 2011 Aug 28;477(7365):495-8. doi: 10.1038/nature10370. Nature. 2011. PMID: 21874019 Free PMC article.
-
Conformational changes at cytoplasmic intersubunit interactions control Kir channel gating.J Biol Chem. 2017 Jun 16;292(24):10087-10096. doi: 10.1074/jbc.M117.785154. Epub 2017 Apr 26. J Biol Chem. 2017. PMID: 28446610 Free PMC article.
-
Hydrogen sulfide inhibits Kir2 and Kir3 channels by decreasing sensitivity to the phospholipid phosphatidylinositol 4,5-bisphosphate (PIP2).J Biol Chem. 2018 Mar 9;293(10):3546-3561. doi: 10.1074/jbc.RA117.001679. Epub 2018 Jan 9. J Biol Chem. 2018. PMID: 29317494 Free PMC article.
-
Phosphoinositide sensitivity of ion channels, a functional perspective.Subcell Biochem. 2012;59:289-333. doi: 10.1007/978-94-007-3015-1_10. Subcell Biochem. 2012. PMID: 22374095 Review.
-
Activation of inwardly rectifying potassium (Kir) channels by phosphatidylinosital-4,5-bisphosphate (PIP2): interaction with other regulatory ligands.Prog Biophys Mol Biol. 2007 Jul;94(3):320-35. doi: 10.1016/j.pbiomolbio.2006.04.001. Epub 2006 Jun 19. Prog Biophys Mol Biol. 2007. PMID: 16837026 Review.
Cited by
-
A Closely Associated Phospholipase C Regulates Cation Channel Function through Phosphoinositide Hydrolysis.J Neurosci. 2018 Aug 29;38(35):7622-7634. doi: 10.1523/JNEUROSCI.0586-18.2018. Epub 2018 Jul 23. J Neurosci. 2018. PMID: 30037836 Free PMC article.
-
Bivalent recognition of fatty acyl-CoA by a human integral membrane palmitoyltransferase.Proc Natl Acad Sci U S A. 2022 Feb 15;119(7):e2022050119. doi: 10.1073/pnas.2022050119. Proc Natl Acad Sci U S A. 2022. PMID: 35140179 Free PMC article.
-
Potassium Channel Gain of Function in Epilepsy: An Unresolved Paradox.Neuroscientist. 2018 Aug;24(4):368-380. doi: 10.1177/1073858418763752. Epub 2018 Mar 15. Neuroscientist. 2018. PMID: 29542386 Free PMC article.
-
Cooperative Gating of a K+ Channel by Unmodified Biological Anionic Lipids Viewed by Solid-State NMR Spectroscopy.J Am Chem Soc. 2024 Feb 21;146(7):4421-4432. doi: 10.1021/jacs.3c09266. Epub 2024 Feb 9. J Am Chem Soc. 2024. PMID: 38334076 Free PMC article.
-
The actions of volatile anesthetics: a new perspective.Acta Crystallogr D Struct Biol. 2018 Dec 1;74(Pt 12):1169-1177. doi: 10.1107/S2059798318004771. Epub 2018 Nov 30. Acta Crystallogr D Struct Biol. 2018. PMID: 30605131 Free PMC article. Review.
References
-
- Hilgemann DW. Local PIP(2) signals: when, where, and how? Pflugers Arch. 2007;455:55–67. - PubMed
-
- Huang C-L. Complex roles of PIP2 in the regulation of ion channels and transporters. Am J Physiol Renal Physiol. 2007;293:F1761–5. - PubMed
-
- Gamper N, Shapiro MS. Regulation of ion transport proteins by membrane phosphoinositides. Nat Rev Neurosci. 2007;8:921–934. - PubMed
-
- Xie L-H, John Sa, Ribalet B, Weiss JN. Activation of inwardly rectifying potassium (Kir) channels by phosphatidylinosital-4,5-bisphosphate (PIP2): interaction with other regulatory ligands. Prog Biophys Mol Biol. 2007;94:320–35. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
