

Translation of 5' leaders is pervasive in genes resistant to eIF2 repression
- PMID: 25621764
- PMCID: PMC4383229
- DOI: 10.7554/eLife.03971
Translation of 5' leaders is pervasive in genes resistant to eIF2 repression
Abstract
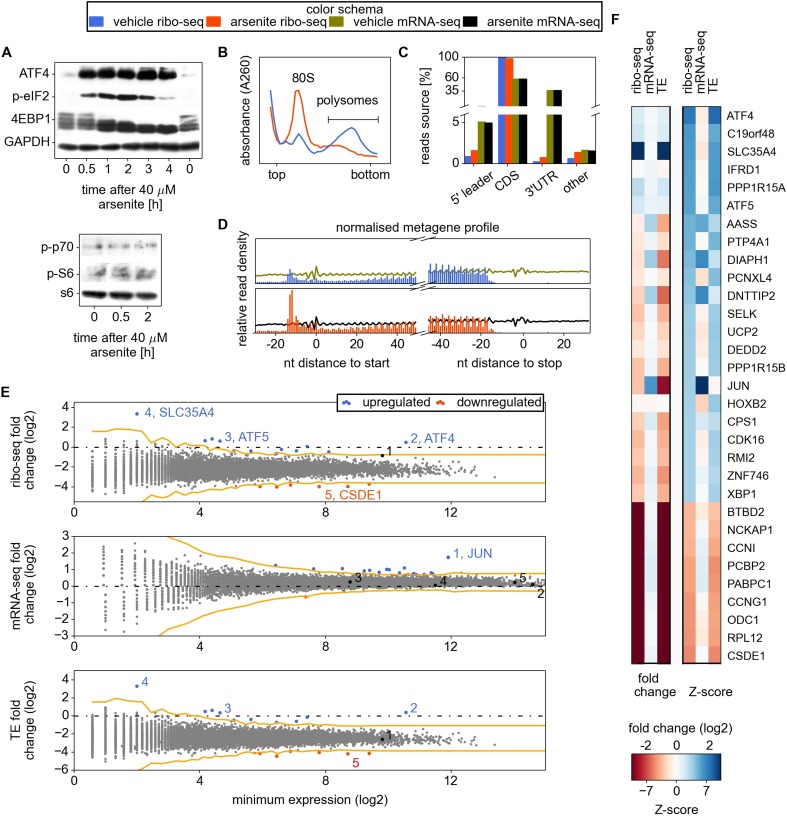
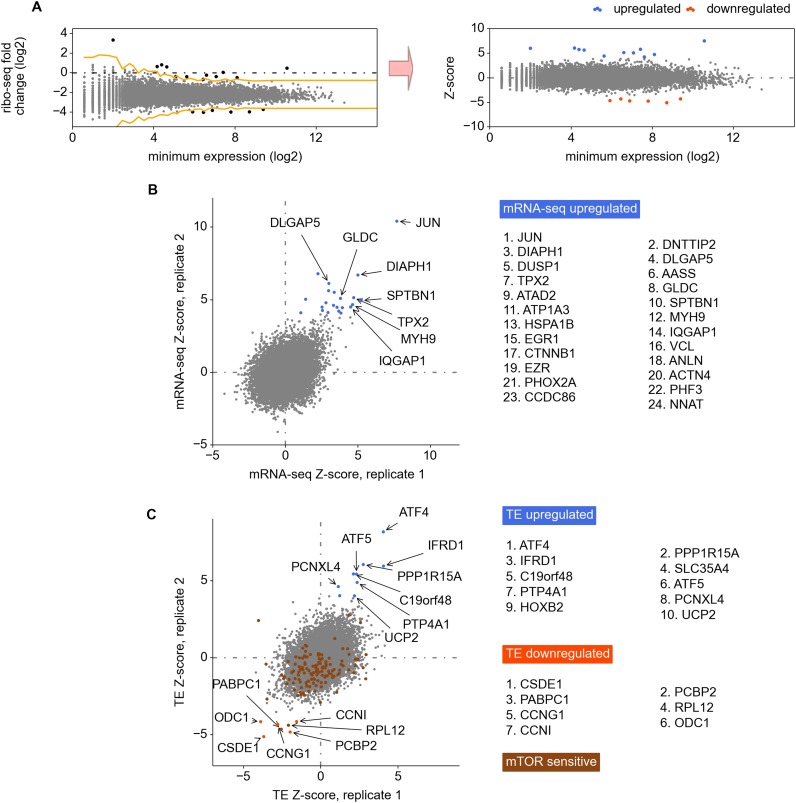
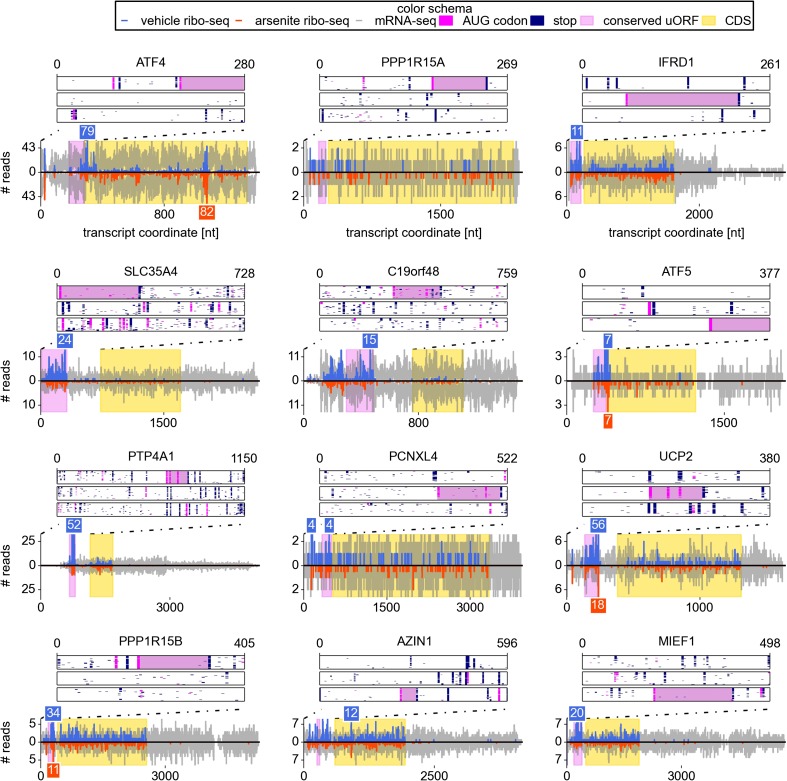
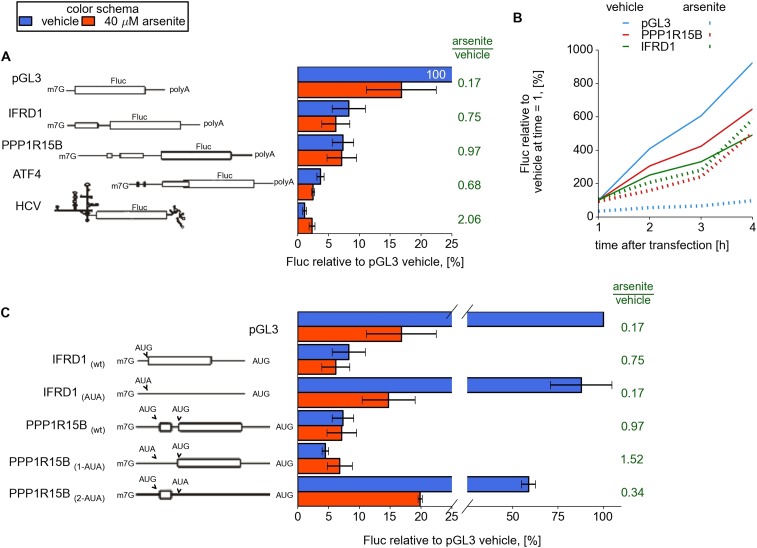
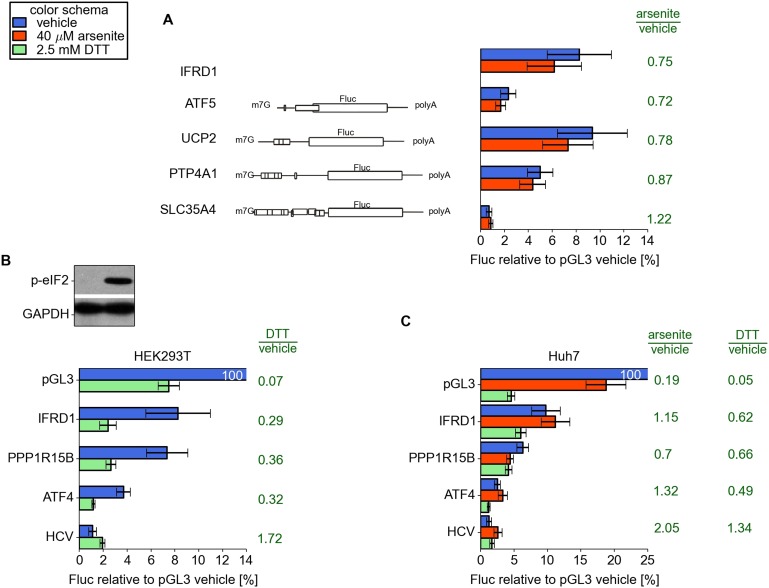
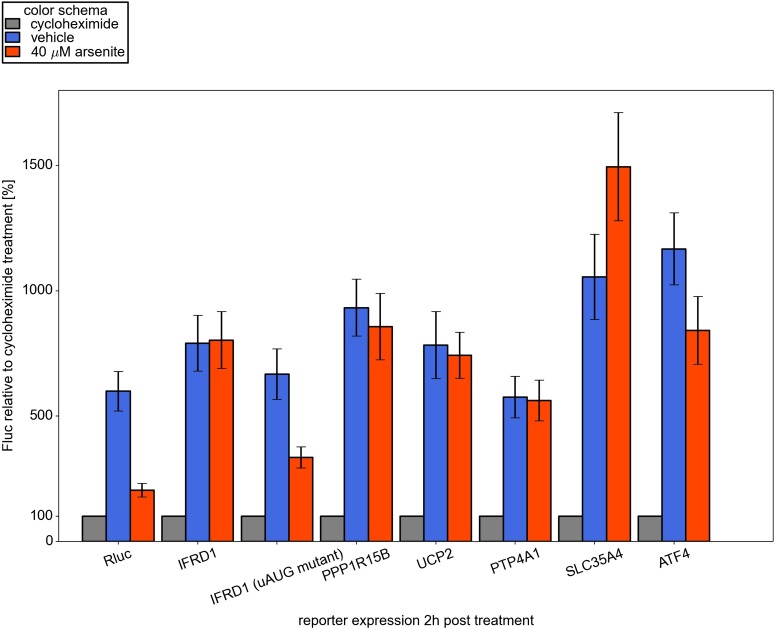
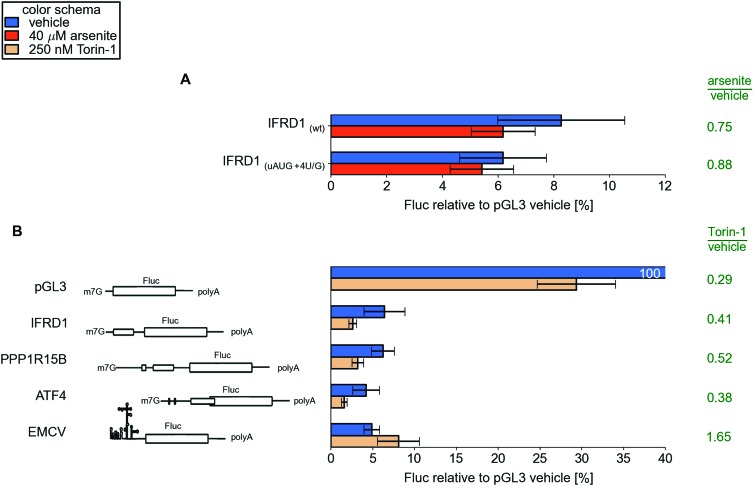
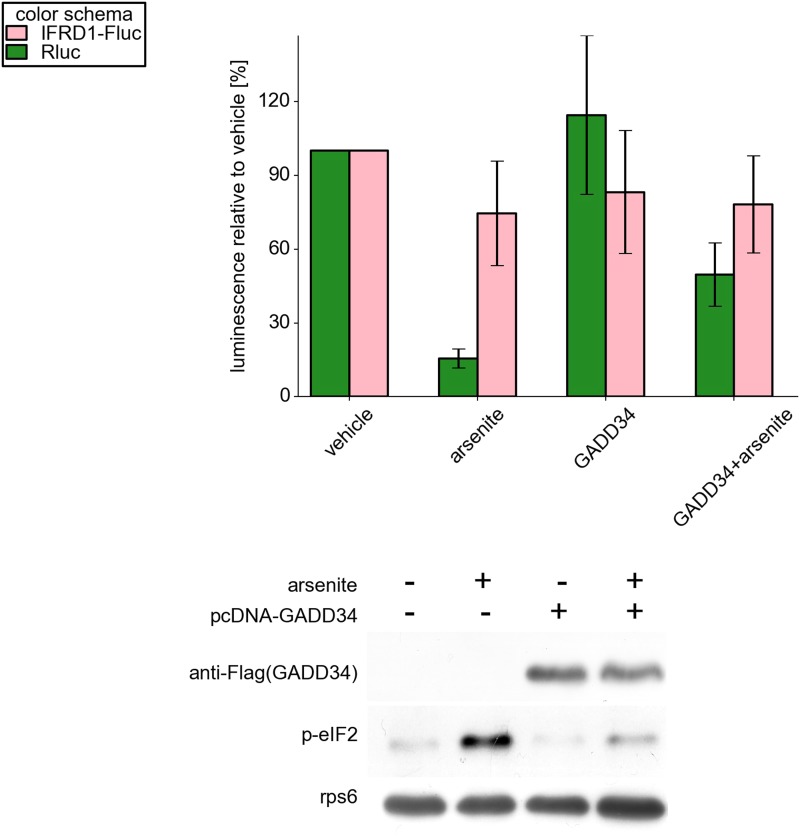
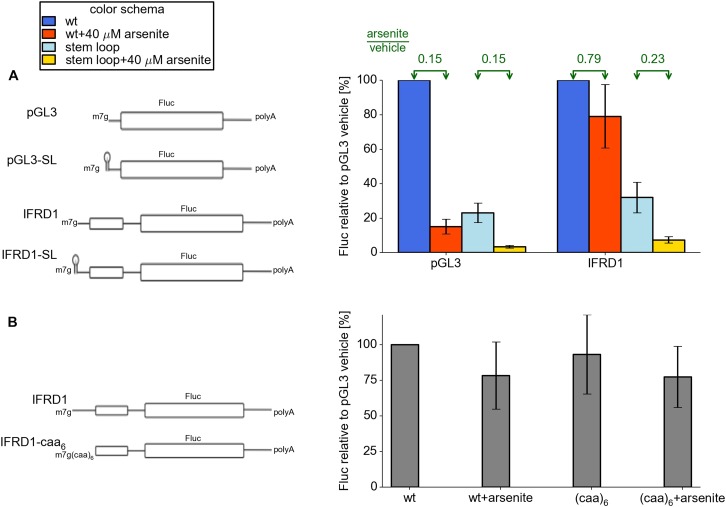
Eukaryotic cells rapidly reduce protein synthesis in response to various stress conditions. This can be achieved by the phosphorylation-mediated inactivation of a key translation initiation factor, eukaryotic initiation factor 2 (eIF2). However, the persistent translation of certain mRNAs is required for deployment of an adequate stress response. We carried out ribosome profiling of cultured human cells under conditions of severe stress induced with sodium arsenite. Although this led to a 5.4-fold general translational repression, the protein coding open reading frames (ORFs) of certain individual mRNAs exhibited resistance to the inhibition. Nearly all resistant transcripts possess at least one efficiently translated upstream open reading frame (uORF) that represses translation of the main coding ORF under normal conditions. Site-specific mutagenesis of two identified stress resistant mRNAs (PPP1R15B and IFRD1) demonstrated that a single uORF is sufficient for eIF2-mediated translation control in both cases. Phylogenetic analysis suggests that at least two regulatory uORFs (namely, in SLC35A4 and MIEF1) encode functional protein products.
Keywords: 5′ leader translation; bicistronic mRNA; biochemistry; eukaryotic initiation factor 2 (eIF2); evolutionary biology; genomics; human; integrated stress response (ISR); ribosome profiling; upstream open reading frame (uORF).
Conflict of interest statement
The authors declare that no competing interests exist.
Figures

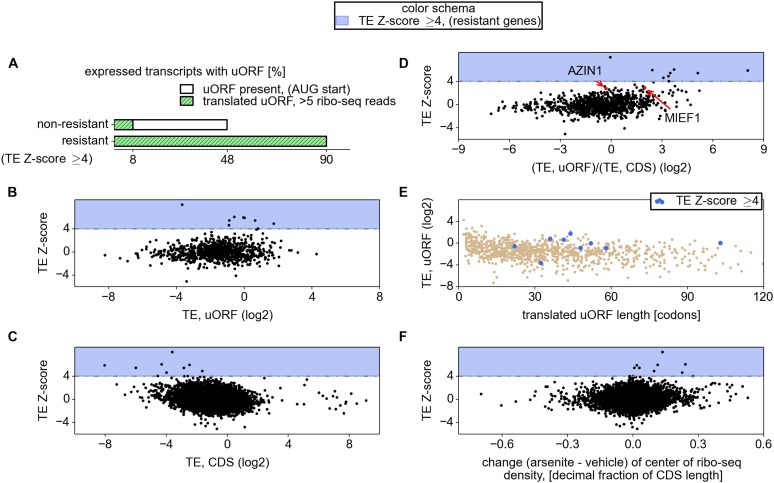
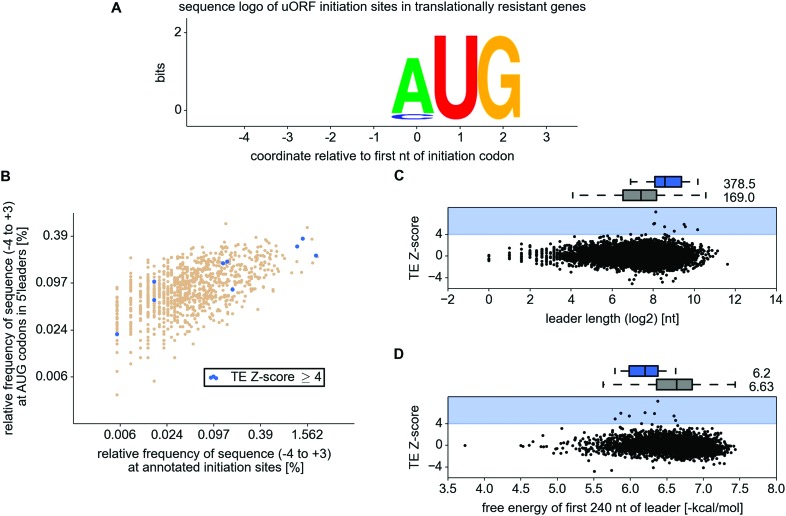
Similar articles
-
TASEP modelling provides a parsimonious explanation for the ability of a single uORF to derepress translation during the integrated stress response.Elife. 2018 Jun 22;7:e32563. doi: 10.7554/eLife.32563. Elife. 2018. PMID: 29932418 Free PMC article.
-
Ribosome Reinitiation Directs Gene-specific Translation and Regulates the Integrated Stress Response.J Biol Chem. 2015 Nov 20;290(47):28257-28271. doi: 10.1074/jbc.M115.693184. Epub 2015 Oct 7. J Biol Chem. 2015. PMID: 26446796 Free PMC article.
-
Phosphorylation of eIF2 facilitates ribosomal bypass of an inhibitory upstream ORF to enhance CHOP translation.J Biol Chem. 2011 Apr 1;286(13):10939-49. doi: 10.1074/jbc.M110.216093. Epub 2011 Feb 1. J Biol Chem. 2011. PMID: 21285359 Free PMC article.
-
Upstream Open Reading Frames Differentially Regulate Gene-specific Translation in the Integrated Stress Response.J Biol Chem. 2016 Aug 12;291(33):16927-35. doi: 10.1074/jbc.R116.733899. Epub 2016 Jun 29. J Biol Chem. 2016. PMID: 27358398 Free PMC article. Review.
-
Translational regulation by uORFs and start codon selection stringency.Genes Dev. 2023 Jun 1;37(11-12):474-489. doi: 10.1101/gad.350752.123. Epub 2023 Jul 11. Genes Dev. 2023. PMID: 37433636 Free PMC article. Review.
Cited by
-
eIF5B drives integrated stress response-dependent translation of PD-L1 in lung cancer.Nat Cancer. 2020 May;1(5):533-545. doi: 10.1038/s43018-020-0056-0. Epub 2020 Apr 20. Nat Cancer. 2020. PMID: 32984844 Free PMC article.
-
Ribonuclease selection for ribosome profiling.Nucleic Acids Res. 2017 Jan 25;45(2):e6. doi: 10.1093/nar/gkw822. Epub 2016 Sep 15. Nucleic Acids Res. 2017. PMID: 27638886 Free PMC article.
-
Ribosome-Profiling Reveals Restricted Post Transcriptional Expression of Antiviral Cytokines and Transcription Factors during SARS-CoV-2 Infection.Int J Mol Sci. 2021 Mar 25;22(7):3392. doi: 10.3390/ijms22073392. Int J Mol Sci. 2021. PMID: 33806254 Free PMC article.
-
Impact of eIF2α phosphorylation on the translational landscape of mouse embryonic stem cells.Cell Rep. 2024 Jan 23;43(1):113615. doi: 10.1016/j.celrep.2023.113615. Epub 2023 Dec 29. Cell Rep. 2024. PMID: 38159280 Free PMC article.
-
Nowhere to hide: unconventional translation yields cryptic peptides for immune surveillance.Immunol Rev. 2016 Jul;272(1):8-16. doi: 10.1111/imr.12434. Immunol Rev. 2016. PMID: 27319338 Free PMC article. Review.
References
-
- Andreev DE, Dmitriev SE, Terenin IM, Shatsky IN. Cap-independent translation initiation of apaf-1 mRNA based on a scanning mechanism is determined by some features of the secondary structure of its 5' untranslated region. Biochemistry Biokhimiia. 2013;78:157–165. doi: 10.1134/S0006297913020041. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
