

Different requirements for Wnt signaling in tongue myogenic subpopulations
- PMID: 25576472
- PMCID: PMC4336158
- DOI: 10.1177/0022034514566030
Different requirements for Wnt signaling in tongue myogenic subpopulations
Abstract
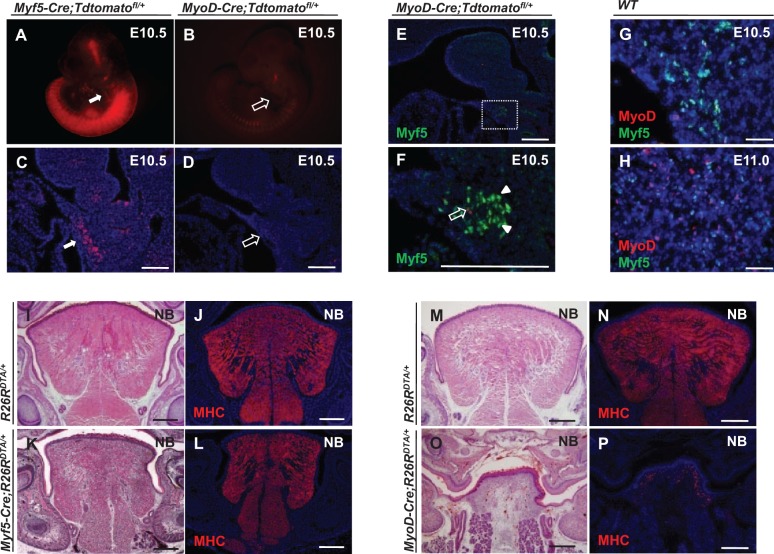
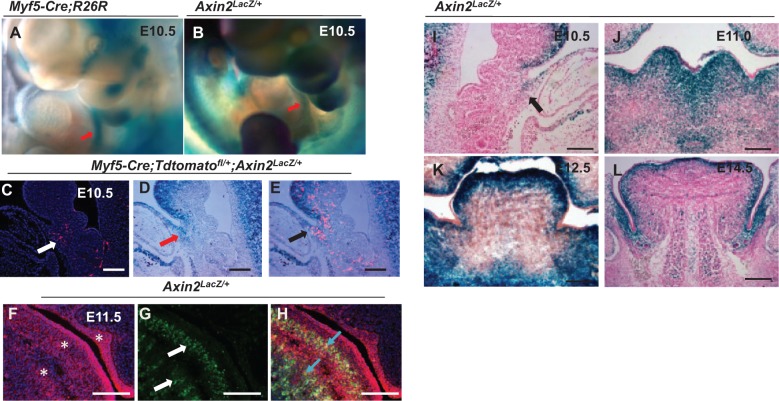
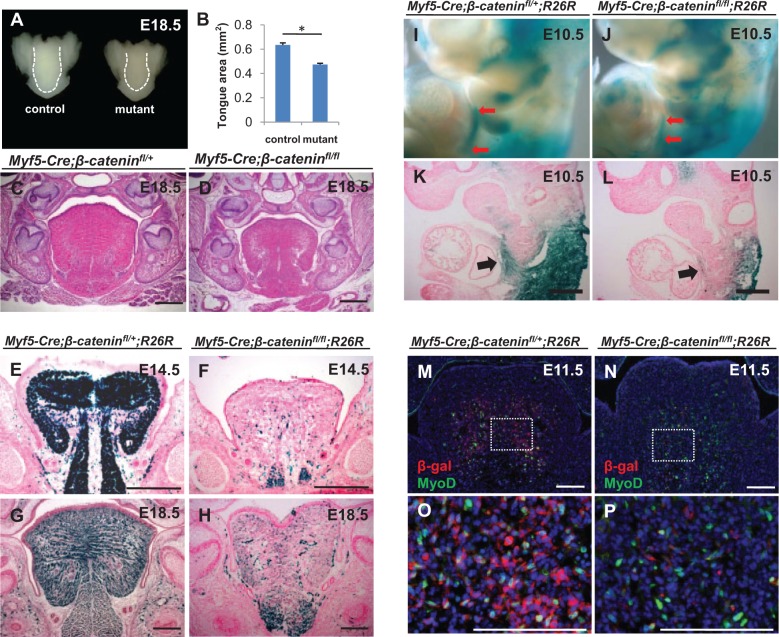
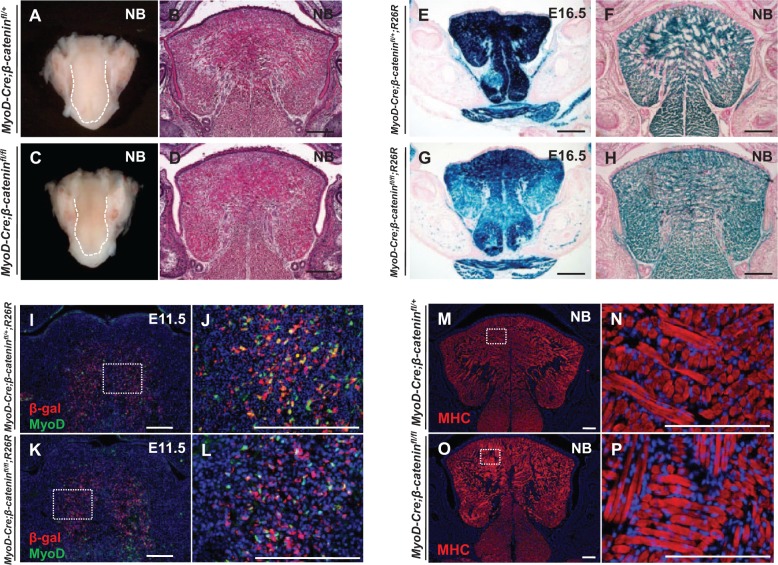
The tongue is a muscular organ that is essential in vertebrates for important functions, such as food intake and communication. Little is known about regulation of myogenic progenitors during tongue development when compared with the limb or trunk region. In this study, we investigated the relationship between different myogenic subpopulations and the function of canonical Wnt signaling in regulating these subpopulations. We found that Myf5- and MyoD-expressing myogenic subpopulations exist during embryonic tongue myogenesis. In the Myf5-expressing myogenic progenitors, there is a cell-autonomous requirement for canonical Wnt signaling for cell migration and differentiation. In contrast, the MyoD-expressing subpopulation does not require canonical Wnt signaling during tongue myogenesis. Taken together, our results demonstrate that canonical Wnt signaling differentially regulates the Myf5- and MyoD-expressing subpopulations during tongue myogenesis.
Keywords: Myf5; MyoD; Wnt/β-catenin; craniofacial muscles; myogenesis; progenitors.
© International & American Associations for Dental Research 2015.
Conflict of interest statement
The authors declare no potential conflicts of interest with respect to the authorship and/or publication of this article.
Figures
Similar articles
-
Different autonomous myogenic cell populations revealed by ablation of Myf5-expressing cells during mouse embryogenesis.Development. 2008 May;135(9):1597-604. doi: 10.1242/dev.019331. Epub 2008 Mar 26. Development. 2008. PMID: 18367555
-
A TGFβ-Smad4-Fgf6 signaling cascade controls myogenic differentiation and myoblast fusion during tongue development.Development. 2012 May;139(9):1640-50. doi: 10.1242/dev.076653. Epub 2012 Mar 21. Development. 2012. PMID: 22438570 Free PMC article.
-
A WNT/beta-catenin signaling activator, R-spondin, plays positive regulatory roles during skeletal myogenesis.J Biol Chem. 2011 Mar 25;286(12):10649-59. doi: 10.1074/jbc.M110.169391. Epub 2011 Jan 20. J Biol Chem. 2011. PMID: 21252233 Free PMC article.
-
Myogenic regulatory factors and the specification of muscle progenitors in vertebrate embryos.Annu Rev Cell Dev Biol. 2002;18:747-83. doi: 10.1146/annurev.cellbio.18.012502.105758. Epub 2002 Apr 2. Annu Rev Cell Dev Biol. 2002. PMID: 12142270 Review.
-
The molecular regulation of muscle stem cell function.Cold Spring Harb Symp Quant Biol. 2008;73:323-31. doi: 10.1101/sqb.2008.73.064. Epub 2009 Mar 27. Cold Spring Harb Symp Quant Biol. 2008. PMID: 19329572 Review.
Cited by
-
Mandible and Tongue Development.Curr Top Dev Biol. 2015;115:31-58. doi: 10.1016/bs.ctdb.2015.07.023. Epub 2015 Oct 1. Curr Top Dev Biol. 2015. PMID: 26589920 Free PMC article. Review.
-
Increased activity of mesenchymal ALK2-BMP signaling causes posteriorly truncated microglossia and disorganization of lingual tissues.Genesis. 2020 Jan;58(1):e23337. doi: 10.1002/dvg.23337. Epub 2019 Sep 30. Genesis. 2020. PMID: 31571391 Free PMC article.
-
WNT/β-catenin signaling plays a crucial role in myoblast fusion through regulation of nephrin expression during development.Development. 2018 Nov 27;145(23):dev168351. doi: 10.1242/dev.168351. Development. 2018. PMID: 30389854 Free PMC article.
-
Dynamic activation of Wnt, Fgf, and Hh signaling during soft palate development.PLoS One. 2019 Oct 15;14(10):e0223879. doi: 10.1371/journal.pone.0223879. eCollection 2019. PLoS One. 2019. PMID: 31613912 Free PMC article.
-
Location, Location, Location: Signals in Muscle Specification.J Dev Biol. 2018 May 18;6(2):11. doi: 10.3390/jdb6020011. J Dev Biol. 2018. PMID: 29783715 Free PMC article. Review.
References
-
- Biressi S, Molinaro M, Cossu G. 2007. Cellular heterogeneity during vertebrate skeletal muscle development. Dev Biol. 308:281–293. - PubMed
-
- Brault V, Moore R, Kutsch S, Ishibashi M, Rowitch DH, McMahon AP, Sommer L, Boussadia O, Kemler R. 2001. Inactivation of the beta-catenin gene by Wnt1-Cre-mediated deletion results in dramatic brain malformation and failure of craniofacial development. Development. 128:1253–1264. - PubMed
-
- Braun T, Gautel M. 2011. Transcriptional mechanisms regulating skeletal muscle differentiation, growth and homeostasis. Nat Rev Mol Cell Biol. 12:349–361. - PubMed
-
- Buckingham M, Rigby PW. 2014. Gene regulatory networks and transcriptional mechanisms that control myogenesis. Dev Cell. 28:225–238. - PubMed
-
- Chai Y, Jiang X, Ito Y, Bringas P, Jr, Han J, Rowitch DH, Soriano P, McMahon AP, Sucov HM. 2000. Fate of the mammalian cranial neural crest during tooth and mandibular morphogenesis. Development. 127:1671–1679. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
