

Natural killer cells can inhibit the transmission of human cytomegalovirus in cell culture by using mechanisms from innate and adaptive immune responses
- PMID: 25540381
- PMCID: PMC4325726
- DOI: 10.1128/JVI.03489-14
Natural killer cells can inhibit the transmission of human cytomegalovirus in cell culture by using mechanisms from innate and adaptive immune responses
Abstract
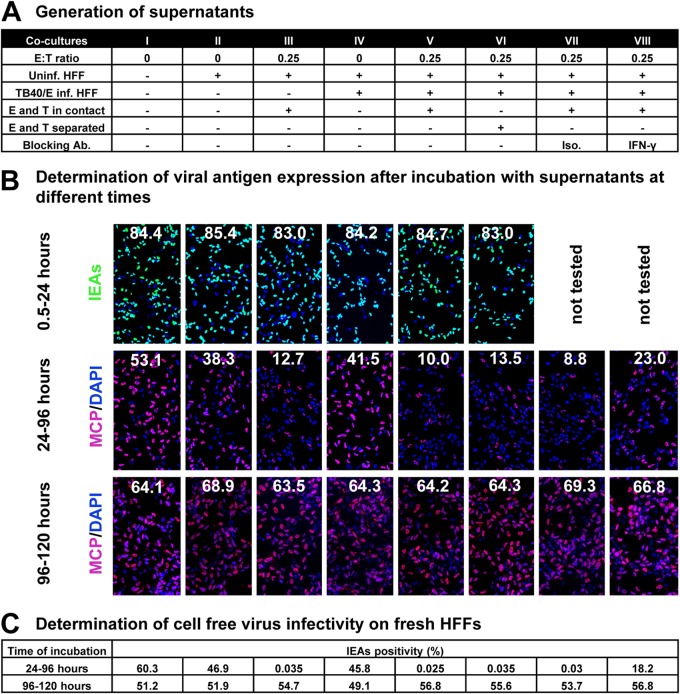
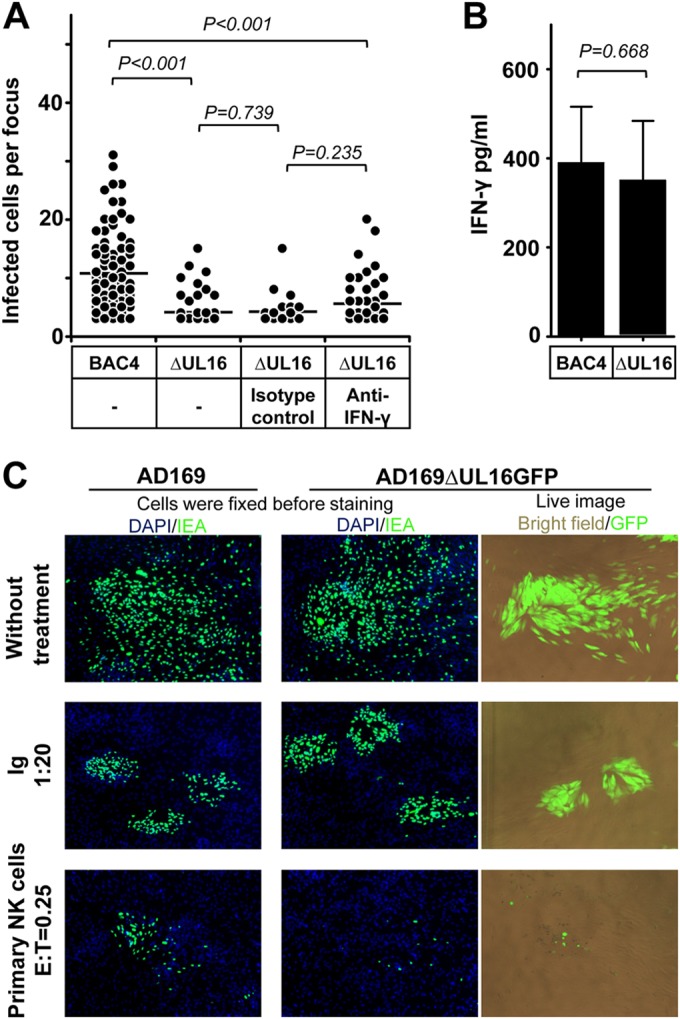
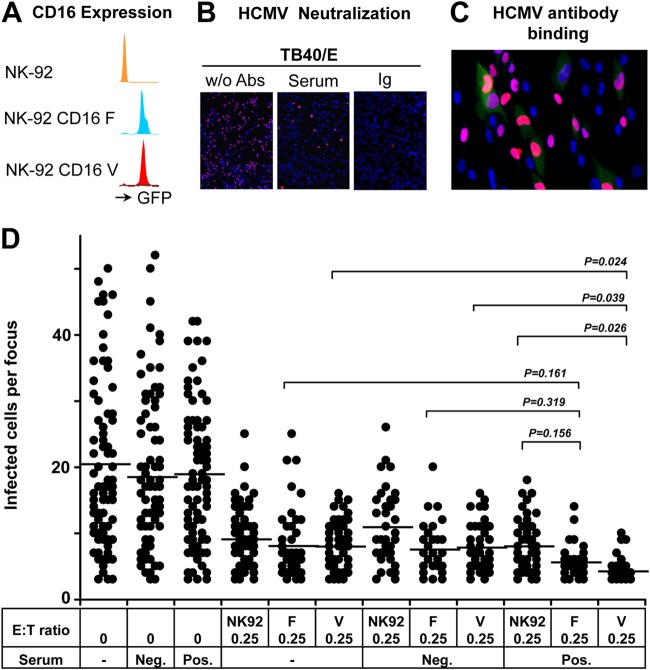
Human cytomegalovirus (HCMV) transmission within the host is important for the pathogenesis of HCMV diseases. Natural killer (NK) cells are well known to provide a first line of host defense against virus infections. However, the role of NK cells in the control of HCMV transmission is still unknown. Here, we provide the first experimental evidence that NK cells can efficiently control HCMV transmission in different cell types. NK cells engage different mechanisms to control the HCMV transmission both via soluble factors and by cell contact. NK cell-produced interferon gamma (IFN-γ) suppresses HCMV production and induces resistance of bystander cells to HCMV infection. The UL16 viral gene contributes to an immune evasion from the NK cell-mediated control of HCMV transmission. Furthermore, the efficacy of the antibody-dependent NK cell-mediated control of HCMV transmission is dependent on a CD16-158V/F polymorphism. Our findings indicate that NK cells may have a clinical relevance in HCMV infection and highlight the need to consider potential therapeutic strategies based on the manipulation of NK cells.
Importance: Human cytomegalovirus (HCMV) infects 40% to 100% of the human population worldwide. After primary infection, mainly in childhood, the virus establishes a lifelong persistence with possible reactivations. Most infections remain asymptomatic; however, HCMV represents a major health problem since it is the most frequent cause of infection-induced birth defects and is responsible for high morbidity and mortality in immunocompromised patients. The immune system normally controls the infection by antibodies and immune effector cells. One type of effector cells are the natural killer (NK) cells, which provide a rapid response to virus-infected cells. NK cells participate in viral clearance by inducing the death of infected cells. NK cells also secrete antiviral cytokines as a consequence of the interaction with an infected cell. In this study, we investigated the mechanisms by which NK cells control HCMV transmission, from the perspectives of immune surveillance and immune evasion.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
Similar articles
-
Interleukin-2 from Adaptive T Cells Enhances Natural Killer Cell Activity against Human Cytomegalovirus-Infected Macrophages.J Virol. 2015 Jun;89(12):6435-41. doi: 10.1128/JVI.00435-15. Epub 2015 Apr 8. J Virol. 2015. PMID: 25855747 Free PMC article.
-
Genetic Variability of Human Cytomegalovirus Clinical Isolates Correlates With Altered Expression of Natural Killer Cell-Activating Ligands and IFN-γ.Front Immunol. 2021 Apr 9;12:532484. doi: 10.3389/fimmu.2021.532484. eCollection 2021. Front Immunol. 2021. PMID: 33897679 Free PMC article.
-
The Human Cytomegalovirus Protein UL148A Downregulates the NK Cell-Activating Ligand MICA To Avoid NK Cell Attack.J Virol. 2018 Aug 16;92(17):e00162-18. doi: 10.1128/JVI.00162-18. Print 2018 Sep 1. J Virol. 2018. PMID: 29950412 Free PMC article.
-
Dendritic cells in cytomegalovirus infection: viral evasion and host countermeasures.APMIS. 2009 May;117(5-6):413-26. doi: 10.1111/j.1600-0463.2009.02449.x. APMIS. 2009. PMID: 19400865 Review.
-
The CD94/NKG2C+ NK-cell subset on the edge of innate and adaptive immunity to human cytomegalovirus infection.Semin Immunol. 2014 Apr;26(2):145-51. doi: 10.1016/j.smim.2014.03.002. Epub 2014 Mar 22. Semin Immunol. 2014. PMID: 24666761 Review.
Cited by
-
Peptide vaccination in the presence of adjuvants in patients after hematopoietic stem cell transplantation with CD4+ T cell reconstitution elicits consistent CD8+ T cell responses.Theranostics. 2017 Apr 10;7(6):1705-1718. doi: 10.7150/thno.18301. eCollection 2017. Theranostics. 2017. PMID: 28529646 Free PMC article.
-
Leukocyte Immunoglobulin-Like Receptor 1-Expressing Human Natural Killer Cell Subsets Differentially Recognize Isolates of Human Cytomegalovirus through the Viral Major Histocompatibility Complex Class I Homolog UL18.J Virol. 2016 Jan 6;90(6):3123-37. doi: 10.1128/JVI.02614-15. J Virol. 2016. PMID: 26739048 Free PMC article.
-
Assessing Anti-HCMV Cell Mediated Immune Responses in Transplant Recipients and Healthy Controls Using a Novel Functional Assay.Front Cell Infect Microbiol. 2020 Jun 26;10:275. doi: 10.3389/fcimb.2020.00275. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 32670891 Free PMC article.
-
Expanded clinical-grade NK cells exhibit stronger effects than primary NK cells against HCMV infection.Cell Mol Immunol. 2023 Aug;20(8):895-907. doi: 10.1038/s41423-023-01046-5. Epub 2023 Jun 9. Cell Mol Immunol. 2023. PMID: 37291236 Free PMC article.
-
New extracellular factors in glioblastoma multiforme development: neurotensin, growth differentiation factor-15, sphingosine-1-phosphate and cytomegalovirus infection.Oncotarget. 2018 Jan 9;9(6):7219-7270. doi: 10.18632/oncotarget.24102. eCollection 2018 Jan 23. Oncotarget. 2018. PMID: 29467963 Free PMC article. Review.
References
-
- Pass RF. 2001. Cytomegalovirus, p 2675–2705 InFields BN, Knipe DM, Howley PM, Griffin DE (ed), Fields virology, 4th ed, vol 2 Lippincott Williams & Wilkins, Philadelphia, PA.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
