

Inositol pyrophosphates regulate RNA polymerase I-mediated rRNA transcription in Saccharomyces cerevisiae
- PMID: 25423617
- PMCID: PMC4325516
- DOI: 10.1042/BJ20140798
Inositol pyrophosphates regulate RNA polymerase I-mediated rRNA transcription in Saccharomyces cerevisiae
Abstract
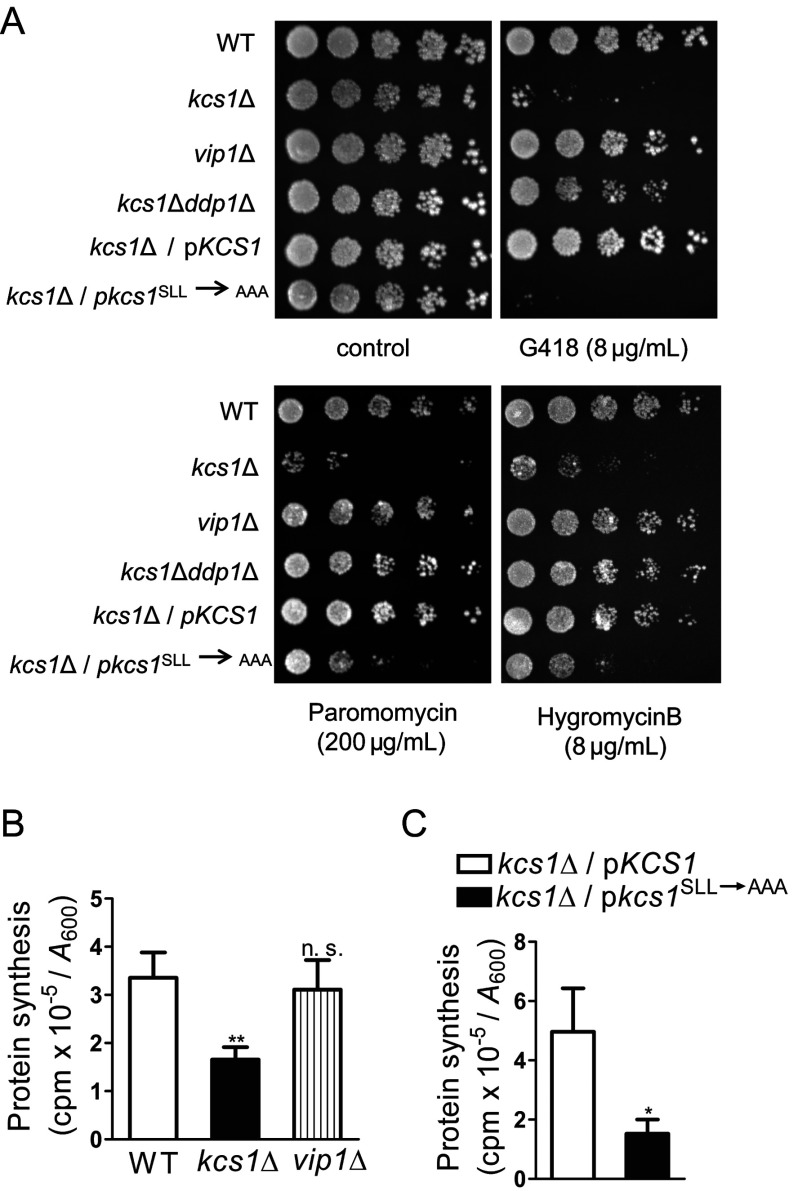
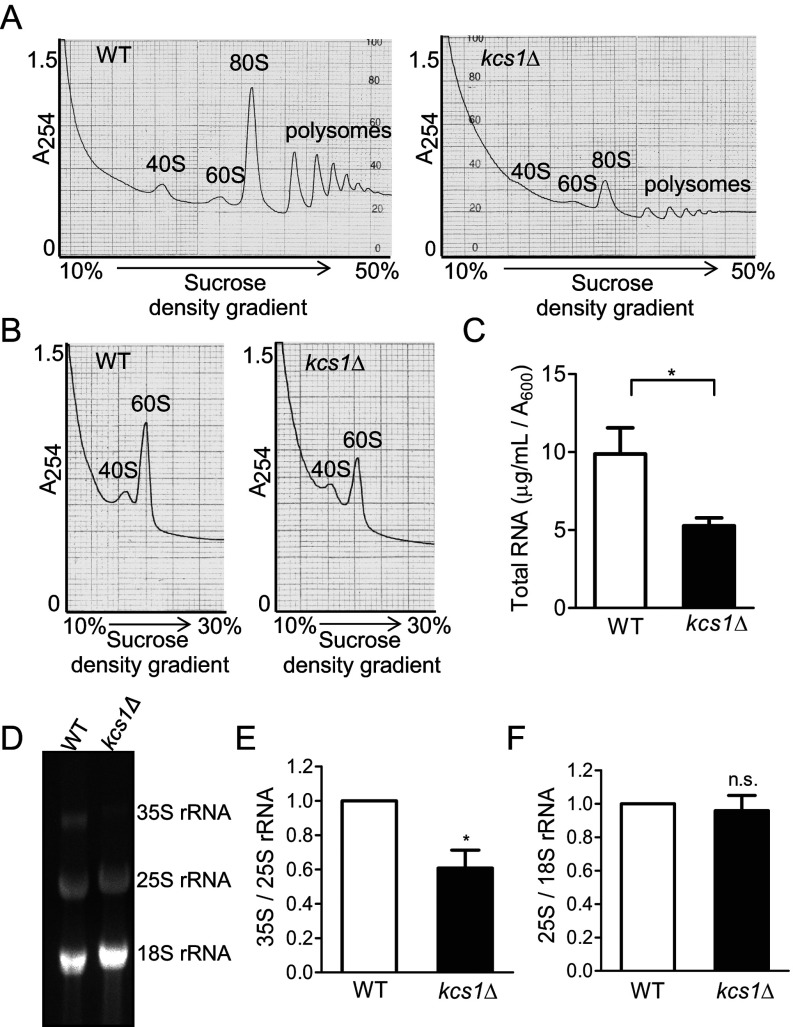
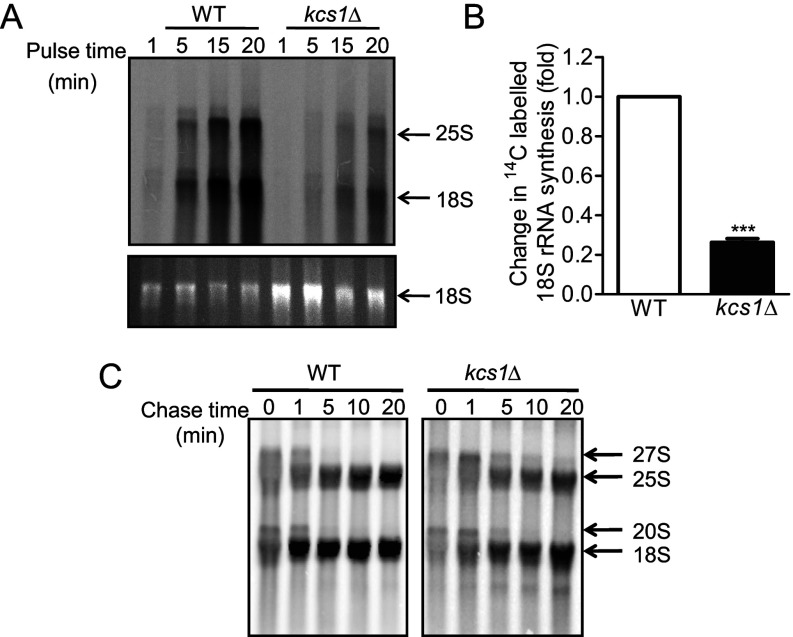
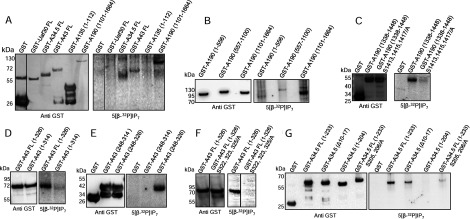
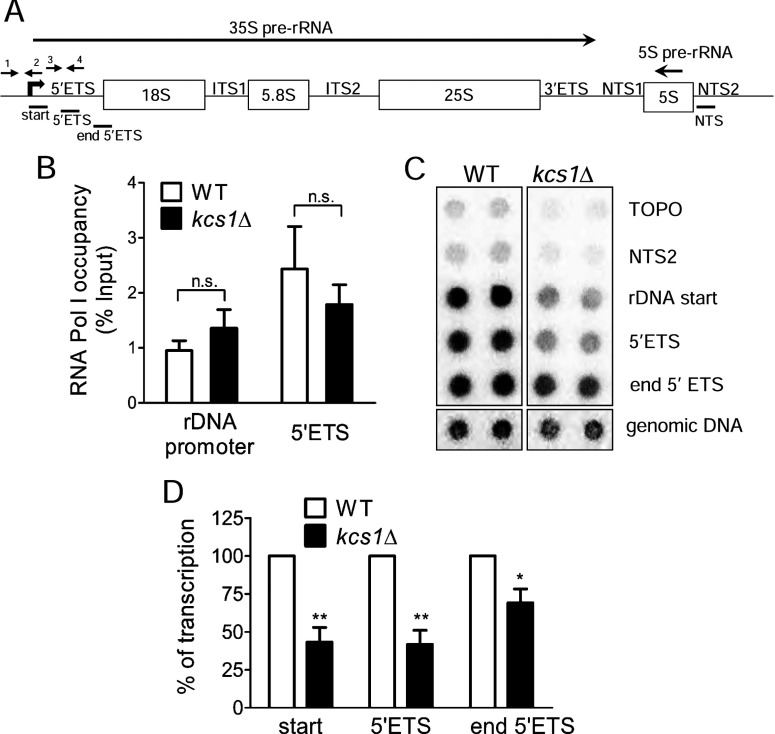
Ribosome biogenesis is an essential cellular process regulated by the metabolic state of a cell. We examined whether inositol pyrophosphates, energy-rich derivatives of inositol that act as metabolic messengers, play a role in ribosome synthesis in the budding yeast, Saccharomyces cerevisiae. Yeast strains lacking the inositol hexakisphosphate (IP6) kinase Kcs1, which is required for the synthesis of inositol pyrophosphates, display increased sensitivity to translation inhibitors and decreased protein synthesis. These phenotypes are reversed on expression of enzymatically active Kcs1, but not on expression of the inactive form. The kcs1Δ yeast cells exhibit reduced levels of ribosome subunits, suggesting that they are defective in ribosome biogenesis. The rate of rRNA synthesis, the first step of ribosome biogenesis, is decreased in kcs1Δ yeast strains, suggesting that RNA polymerase I (Pol I) activity may be reduced in these cells. We determined that the Pol I subunits, A190, A43 and A34.5, can accept a β-phosphate moiety from inositol pyrophosphates to undergo serine pyrophosphorylation. Although there is impaired rRNA synthesis in kcs1Δ yeast cells, we did not find any defect in recruitment of Pol I on rDNA, but observed that the rate of transcription elongation was compromised. Taken together, our findings highlight inositol pyrophosphates as novel regulators of rRNA transcription.
Figures
Similar articles
-
Inositol pyrophosphates modulate S phase progression after pheromone-induced arrest in Saccharomyces cerevisiae.J Biol Chem. 2013 Jan 18;288(3):1717-25. doi: 10.1074/jbc.M112.412288. Epub 2012 Nov 24. J Biol Chem. 2013. PMID: 23179856 Free PMC article.
-
Regulation of inositol metabolism is fine-tuned by inositol pyrophosphates in Saccharomyces cerevisiae.J Biol Chem. 2013 Aug 23;288(34):24898-908. doi: 10.1074/jbc.M113.493353. Epub 2013 Jul 2. J Biol Chem. 2013. PMID: 23824185 Free PMC article.
-
Inositol pyrophosphates modulate hydrogen peroxide signalling.Biochem J. 2009 Sep 14;423(1):109-18. doi: 10.1042/BJ20090241. Biochem J. 2009. PMID: 19614566
-
Kcs1 and Vip1: The Key Enzymes behind Inositol Pyrophosphate Signaling in Saccharomyces cerevisiae.Biomolecules. 2024 Jan 26;14(2):152. doi: 10.3390/biom14020152. Biomolecules. 2024. PMID: 38397389 Free PMC article. Review.
-
Features of yeast RNA polymerase I with special consideration of the lobe binding subunits.Biol Chem. 2023 Oct 13;404(11-12):979-1002. doi: 10.1515/hsz-2023-0184. Print 2023 Oct 26. Biol Chem. 2023. PMID: 37823775 Review.
Cited by
-
The emerging roles of inositol pyrophosphates in eukaryotic cell physiology.J Biosci. 2015 Sep;40(3):593-605. doi: 10.1007/s12038-015-9549-x. J Biosci. 2015. PMID: 26333405 Review.
-
The inositol pyrophosphate pathway in health and diseases.Biol Rev Camb Philos Soc. 2018 May;93(2):1203-1227. doi: 10.1111/brv.12392. Epub 2017 Dec 27. Biol Rev Camb Philos Soc. 2018. PMID: 29282838 Free PMC article. Review.
-
Interaction with IP6K1 supports pyrophosphorylation of substrate proteins by the inositol pyrophosphate 5-InsP7.Biosci Rep. 2024 Oct 30;44(10):BSR20240792. doi: 10.1042/BSR20240792. Biosci Rep. 2024. PMID: 39230924 Free PMC article.
-
Inositol polyphosphates intersect with signaling and metabolic networks via two distinct mechanisms.Proc Natl Acad Sci U S A. 2016 Nov 1;113(44):E6757-E6765. doi: 10.1073/pnas.1606853113. Epub 2016 Oct 19. Proc Natl Acad Sci U S A. 2016. PMID: 27791083 Free PMC article.
-
Inositol Hexakisphosphate Kinase 3 Regulates Metabolism and Lifespan in Mice.Sci Rep. 2016 Aug 31;6:32072. doi: 10.1038/srep32072. Sci Rep. 2016. PMID: 27577108 Free PMC article.
References
-
- Nomura M., Nogi Y., Oakes M. Transcription of rDNA in the yeast Saccharomyces cerevisiae. In: Olson M. O. J., editor. The Nucleolus. Georgetown, TX: Landes Bioscience; 2004. pp. 128–153.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases