

Hibiscus chlorotic ringspot virus coat protein is essential for cell-to-cell and long-distance movement but not for viral RNA replication
- PMID: 25402344
- PMCID: PMC4234673
- DOI: 10.1371/journal.pone.0113347
Hibiscus chlorotic ringspot virus coat protein is essential for cell-to-cell and long-distance movement but not for viral RNA replication
Abstract
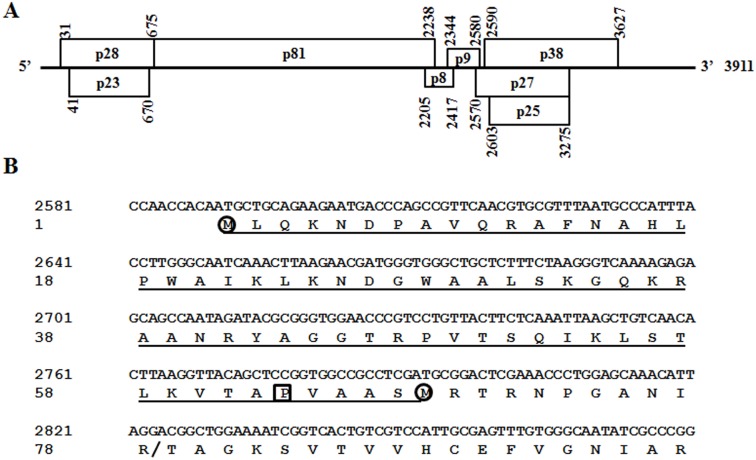
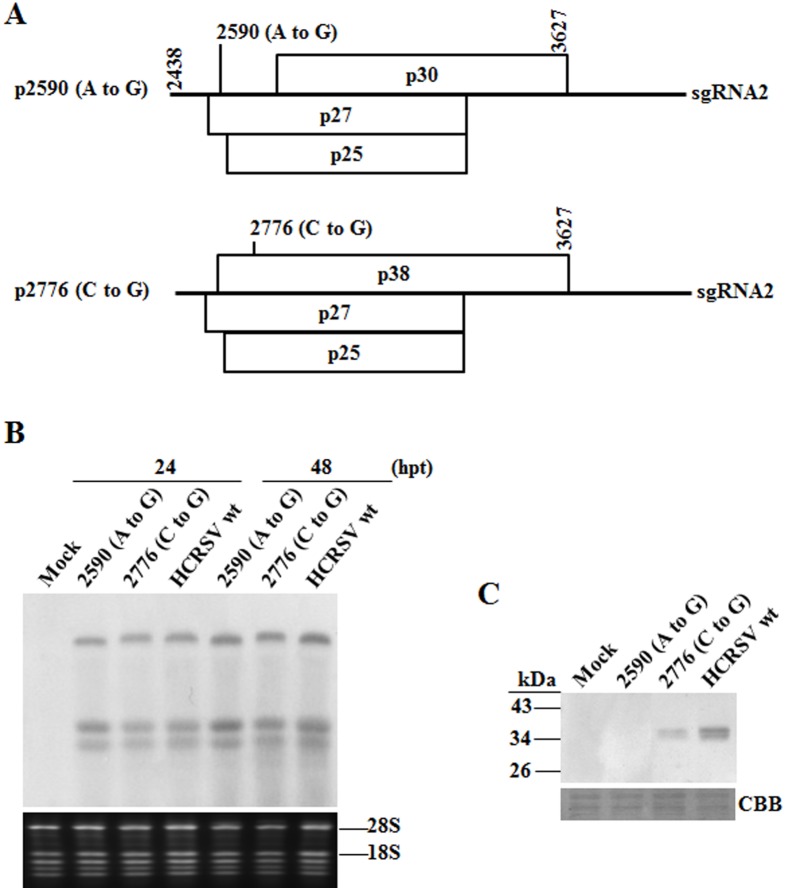
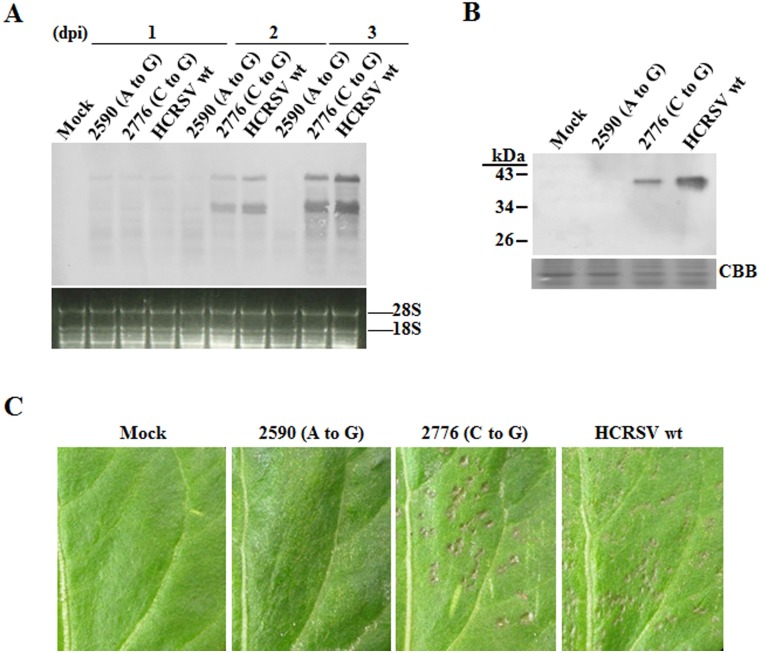
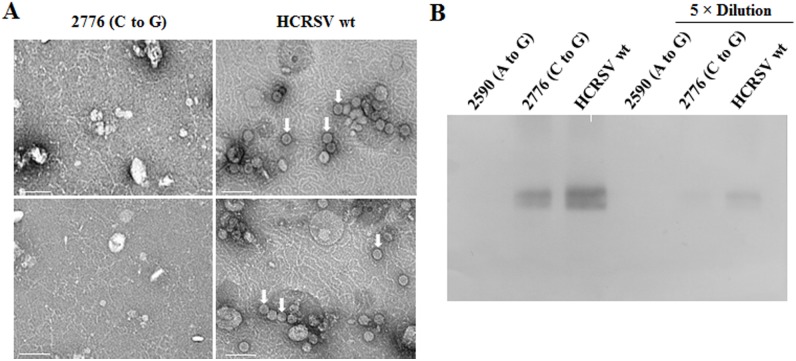
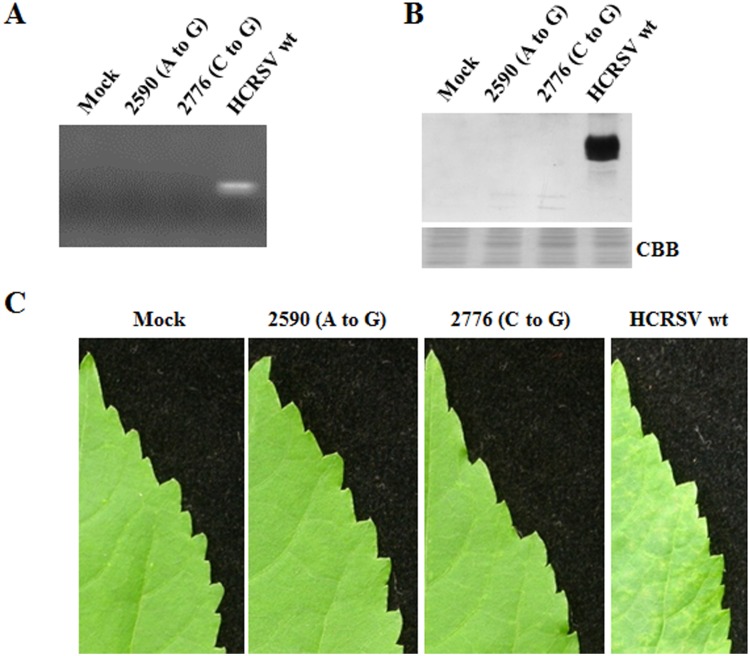
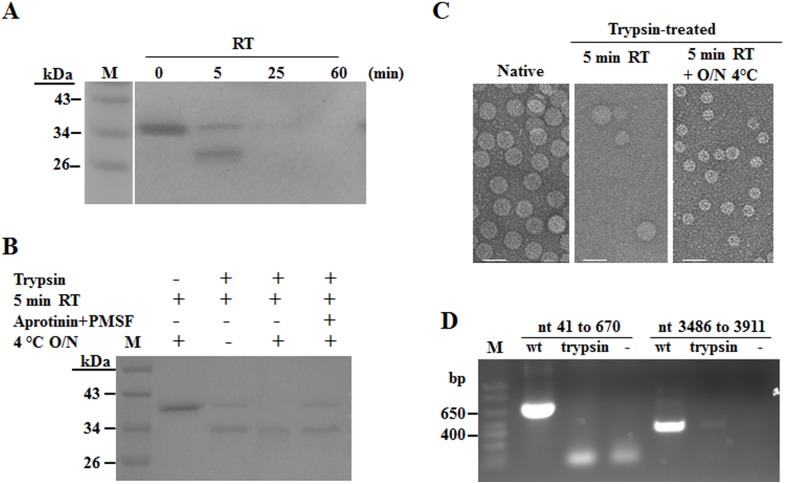
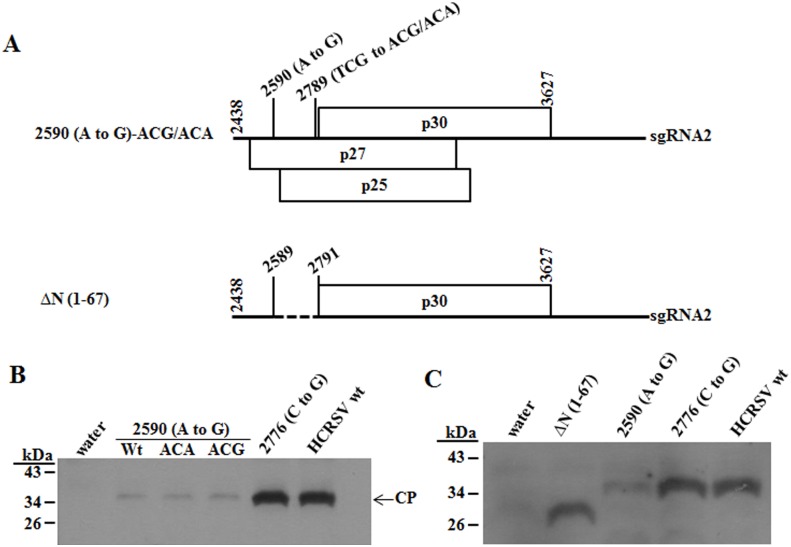
Hibiscus chlorotic ringspot virus (HCRSV) is a member of the genus Carmovirus in the family Tombusviridae. In order to study its coat protein (CP) functions on virus replication and movement in kenaf (Hibiscus cannabinus L.), two HCRSV mutants, designated as p2590 (A to G) in which the first start codon ATG was replaced with GTG and p2776 (C to G) in which proline 63 was replaced with alanine, were constructed. In vitro transcripts of p2590 (A to G) were able to replicate to a similar level as wild type without CP expression in kenaf protoplasts. However, its cell-to-cell movement was not detected in the inoculated kenaf cotyledons. Structurally the proline 63 in subunit C acts as a kink for β-annulus formation during virion assembly. Progeny of transcripts derived from p2776 (C to G) was able to move from cell-to-cell in inoculated cotyledons but its long-distance movement was not detected. Virions were not observed in partially purified mutant virus samples isolated from 2776 (C to G) inoculated cotyledons. Removal of the N-terminal 77 amino acids of HCRSV CP by trypsin digestion of purified wild type HCRSV virions resulted in only T = 1 empty virus-like particles. Taken together, HCRSV CP is dispensable for viral RNA replication but essential for cell-to-cell movement, and virion is required for the virus systemic movement. The proline 63 is crucial for HCRSV virion assembly in kenaf plants and the N-terminal 77 amino acids including the β-annulus domain is required in T = 3 assembly in vitro.
Conflict of interest statement
Figures
Similar articles
-
Hibiscus chlorotic ringspot virus p27 and its isoforms affect symptom expression and potentiate virus movement in kenaf (Hibiscus cannabinus L.).Mol Plant Microbe Interact. 2006 Sep;19(9):948-57. doi: 10.1094/MPMI-19-0948. Mol Plant Microbe Interact. 2006. PMID: 16941899
-
Hibiscus chlorotic ringspot virus upregulates plant sulfite oxidase transcripts and increases sulfate levels in kenaf (Hibiscus cannabinus L.).J Gen Virol. 2009 Dec;90(Pt 12):3042-3050. doi: 10.1099/vir.0.012112-0. Epub 2009 Sep 2. J Gen Virol. 2009. PMID: 19726610
-
Host-induced avirulence of hibiscus chlorotic ringspot virus mutants correlates with reduced gene-silencing suppression activity.J Gen Virol. 2006 Feb;87(Pt 2):451-459. doi: 10.1099/vir.0.81578-0. J Gen Virol. 2006. PMID: 16432034
-
Basic amino acid mutations in the nuclear localization signal of hibiscus chlorotic ringspot virus p23 inhibit virus long distance movement.PLoS One. 2013 Sep 3;8(9):e74000. doi: 10.1371/journal.pone.0074000. eCollection 2013. PLoS One. 2013. PMID: 24019944 Free PMC article.
-
Viral and nonviral elements in potexvirus replication and movement and in antiviral responses.Adv Virus Res. 2013;87:75-112. doi: 10.1016/B978-0-12-407698-3.00003-X. Adv Virus Res. 2013. PMID: 23809921 Review.
Cited by
-
Characterization of Hibiscus Chlorotic Ringspot Virus-Derived vsiRNAs from Infected Hibiscus rosa-sinensis in China.Plant Pathol J. 2024 Oct;40(5):415-424. doi: 10.5423/PPJ.OA.06.2024.0090. Epub 2024 Oct 1. Plant Pathol J. 2024. PMID: 39397297 Free PMC article.
-
The atomic structures of shrimp nodaviruses reveal new dimeric spike structures and particle polymorphism.Commun Biol. 2019 Feb 20;2:72. doi: 10.1038/s42003-019-0311-z. eCollection 2019. Commun Biol. 2019. PMID: 30820467 Free PMC article.
-
Phosphorylation of Beet black scorch virus coat protein by PKA is required for assembly and stability of virus particles.Sci Rep. 2015 Jun 25;5:11585. doi: 10.1038/srep11585. Sci Rep. 2015. PMID: 26108567 Free PMC article.
References
-
- Brunt AA, Crabtree K, Dallwitz MJ, Gibbs AJ, Watson L, et al... (1996) Viruses of plant: Descriptions and lists from the VIDE database. Wallingford, UK: CAB International.
-
- Lommel SA, Martelli GP, Russo M (2000) Family Tombusviridae. In: Van Regenmortel MHV, Fauquet CM, Bishop DHL, Carstens EB, Estes MK et al..., editors. Virus Taxonomy: Seventh Report of the International Committee on the Taxonomy of Viruses. International Union of Microbiological Societies: Elsevier Science & Technology Books. 791–795.
-
- Jones DR, Behncken GM (1980) Hibiscus chlorotic ringspot, a widespread virus disease in the ornamental Hibiscus rosa-sinensis. Aust Plant Pathol 9: 4–5.
-
- Waterworth HE (1980) Hibiscus chlorotic ringspot virus. Descriptions of plant viruses. Warwick, UK: Association of Applied Biologists.
-
- Brunt AA, Spence NJ (2000) The natural occurrence of Hibiscus chlorotic ringspot virus (Carmovirus; Tombusviridae) in aibika or bele (Abelmoschus manihot) in some South Pacific Island countries. Plant Pathol 49: 798.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
