

Transcriptional silencing by polycomb-group proteins
- PMID: 25367972
- PMCID: PMC4413232
- DOI: 10.1101/cshperspect.a019331
Transcriptional silencing by polycomb-group proteins
Abstract
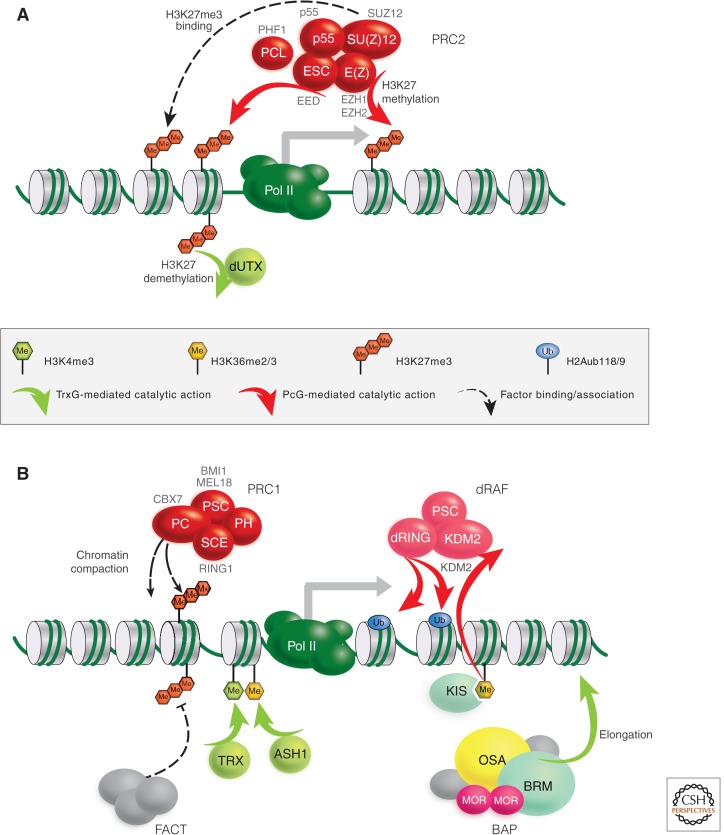
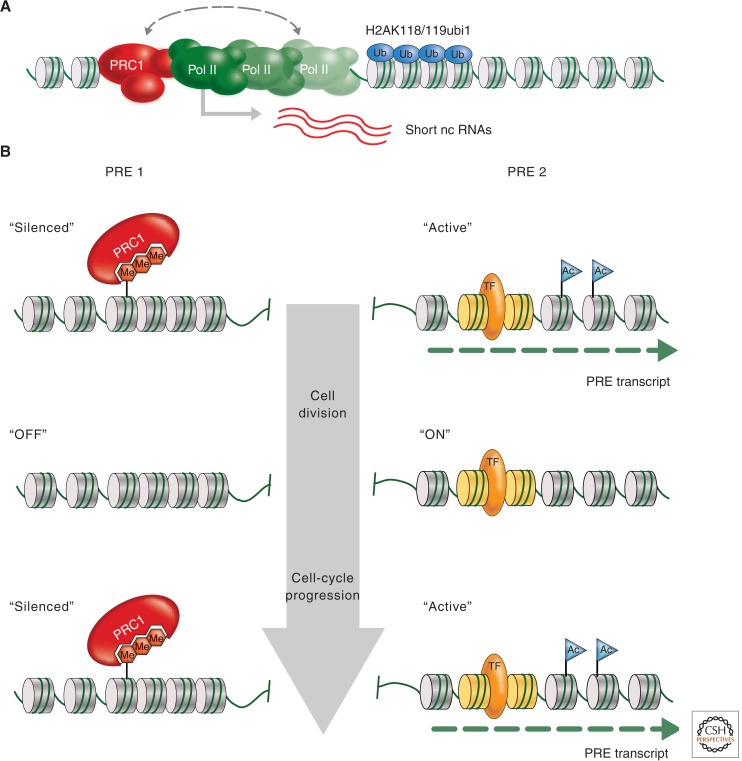
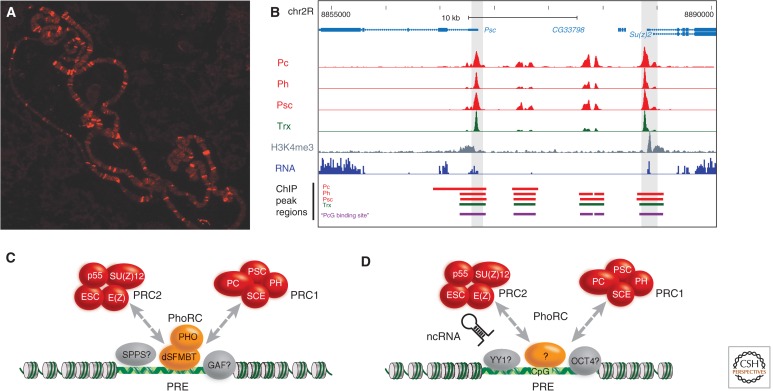
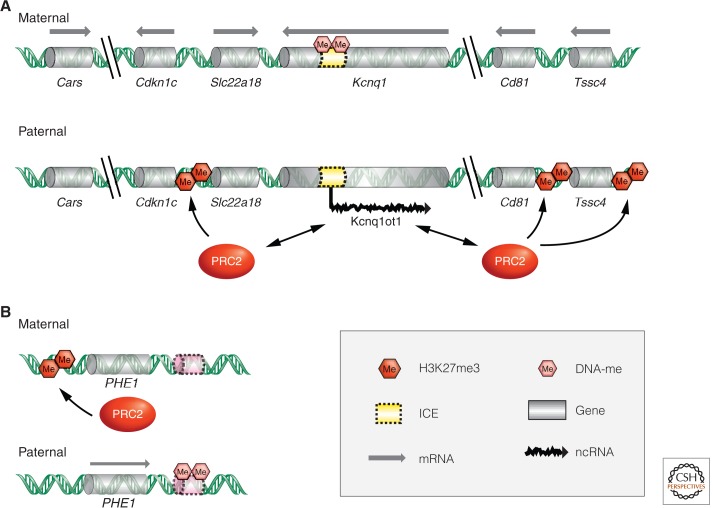
Polycomb-group (PcG) genes encode chromatin proteins involved in stable and heritable transcriptional silencing. PcG proteins participate in distinct multimeric complexes that deposit, or bind to, specific histone modifications (e.g., H3K27me3 and H2AK119ub1) to prevent gene activation and maintain repressed chromatin domains. PcG proteins are evolutionary conserved and play a role in processes ranging from vernalization and seed development in plants, over X-chromosome inactivation in mammals, to the maintenance of stem cell identity. PcG silencing is medically relevant as it is often observed in human disorders, including cancer, and tissue regeneration, which involve the reprogramming of PcG-controlled target genes.
Copyright © 2014 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
Similar articles
-
Concise review: roles of polycomb group proteins in development and disease: a stem cell perspective.Stem Cells. 2007 Oct;25(10):2498-510. doi: 10.1634/stemcells.2006-0608. Epub 2007 Jun 28. Stem Cells. 2007. PMID: 17600113 Review.
-
Polycomb group protein-mediated histone modifications during cell differentiation.Epigenomics. 2015;7(1):75-84. doi: 10.2217/epi.14.61. Epigenomics. 2015. PMID: 25687468 Review.
-
Polycomb silencing: from linear chromatin domains to 3D chromosome folding.Curr Opin Genet Dev. 2014 Apr;25:30-7. doi: 10.1016/j.gde.2013.11.016. Epub 2014 Jan 14. Curr Opin Genet Dev. 2014. PMID: 24434548 Review.
-
CHD3 proteins and polycomb group proteins antagonistically determine cell identity in Arabidopsis.PLoS Genet. 2009 Aug;5(8):e1000605. doi: 10.1371/journal.pgen.1000605. Epub 2009 Aug 14. PLoS Genet. 2009. PMID: 19680533 Free PMC article.
-
Polycomb group proteins and their roles in regulating stem cell development.Zhongguo Yi Xue Ke Xue Yuan Xue Bao. 2012 Jun;34(3):281-5. doi: 10.3881/j.issn.1000-503X.2012.03.019. Zhongguo Yi Xue Ke Xue Yuan Xue Bao. 2012. PMID: 22776663 Review.
Cited by
-
Dosage compensation in mammals.Cold Spring Harb Perspect Biol. 2015 Mar 2;7(3):a019406. doi: 10.1101/cshperspect.a019406. Cold Spring Harb Perspect Biol. 2015. PMID: 25731764 Free PMC article. Review.
-
Age reprogramming: cell rejuvenation by partial reprogramming.Development. 2022 Nov 15;149(22):dev200755. doi: 10.1242/dev.200755. Epub 2022 Nov 16. Development. 2022. PMID: 36383700 Free PMC article.
-
LINC02678 as a Novel Prognostic Marker Promotes Aggressive Non-small-cell Lung Cancer.Front Cell Dev Biol. 2021 May 28;9:686975. doi: 10.3389/fcell.2021.686975. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34124072 Free PMC article.
-
Developmental control of transcriptional and proliferative potency during the evolutionary emergence of animals.Dev Dyn. 2015 Oct;244(10):1193-201. doi: 10.1002/dvdy.24305. Epub 2015 Aug 4. Dev Dyn. 2015. PMID: 26173445 Free PMC article.
-
Transcriptional Stress Memory and Transgenerational Inheritance of Drought Tolerance in Plants.Int J Mol Sci. 2022 Oct 26;23(21):12918. doi: 10.3390/ijms232112918. Int J Mol Sci. 2022. PMID: 36361708 Free PMC article. Review.
References
-
- Agger K, Cloos PA, Christensen J, Pasini D, Rose S, Rappsilber J, Issaeva I, Canaani E, Salcini AE, Helin K 2007. UTX and JMJD3 are histone H3K27 demethylases involved in HOX gene regulation and development. Nature 449: 731–734. - PubMed
-
- Allis CD, Jenuwein T, Reinberg D 2014. Overview and concepts. Cold Spring Harb Perspect Biol 10.1101/cshperspect.a018739. - DOI
-
- Alvarez-Venegas R, Pien S, Sadder M, Witmer X, Grossniklaus U, Avramova Z 2003. ATX-1, an Arabidopsis homolog of trithorax, activates flower homeotic genes. Curr Biol 13: 627–637. - PubMed
-
- Bantignies F, Cavalli G 2011. Polycomb group proteins: Repression in 3D. Trends Genet 27: 454–474. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases