

Structure and immune recognition of trimeric pre-fusion HIV-1 Env
- PMID: 25296255
- PMCID: PMC4348022
- DOI: 10.1038/nature13808
Structure and immune recognition of trimeric pre-fusion HIV-1 Env
Abstract
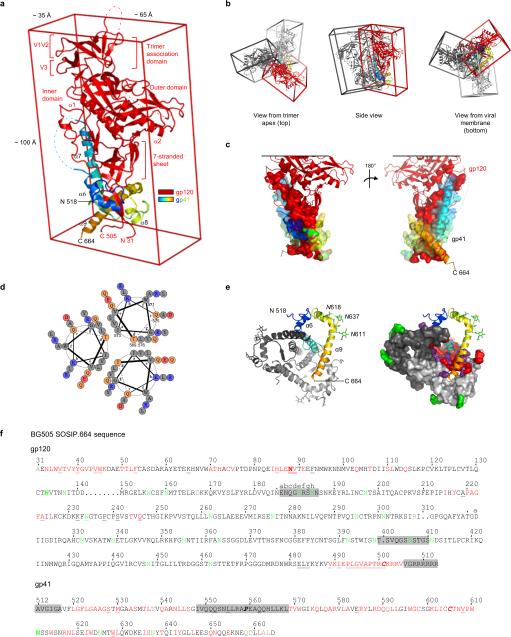
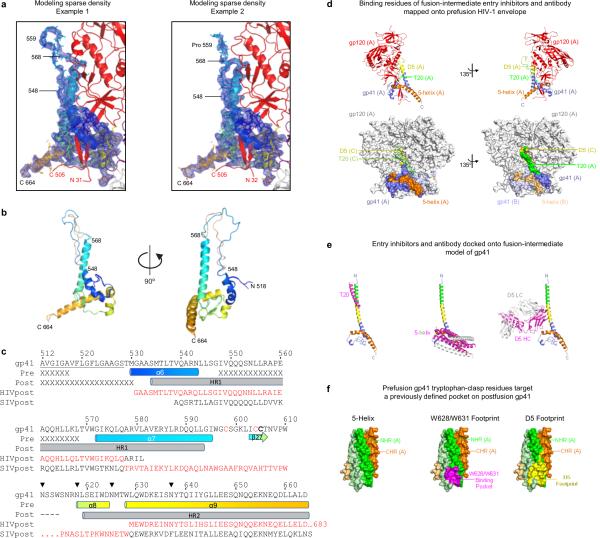
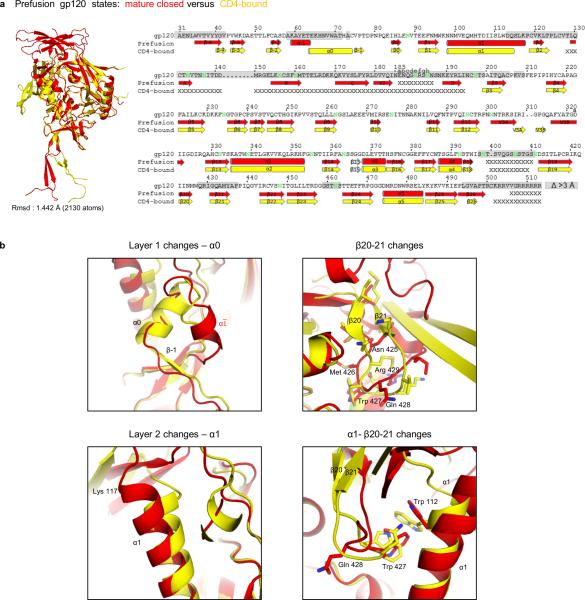
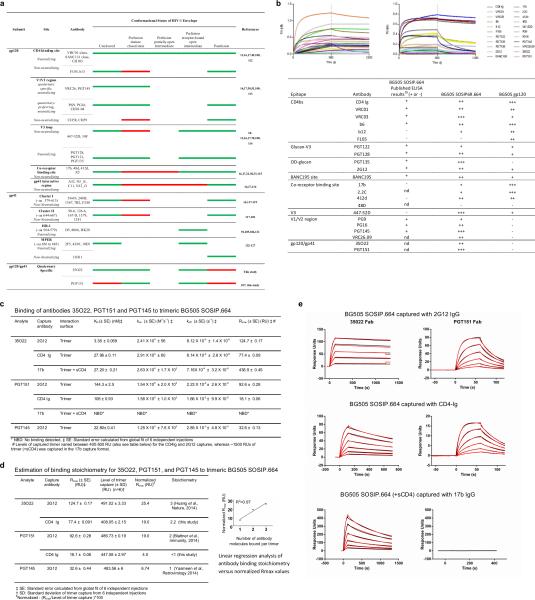
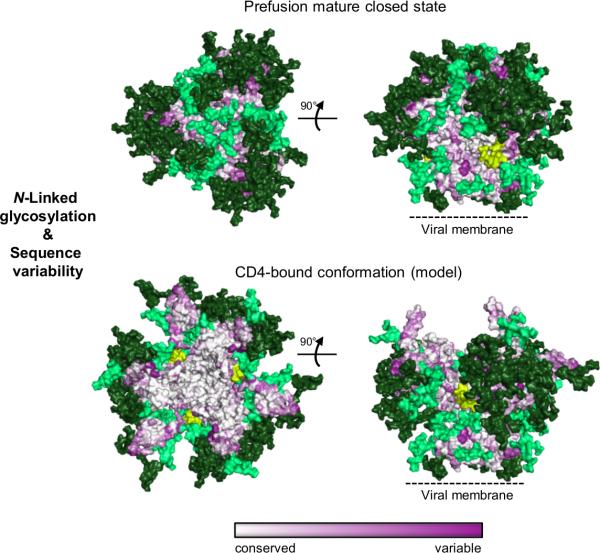
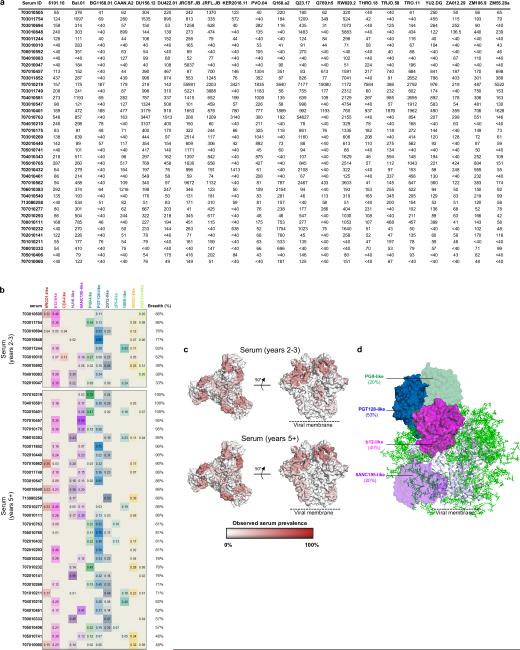
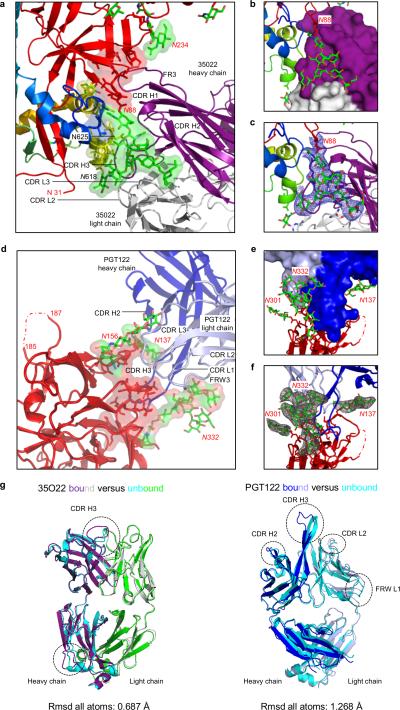
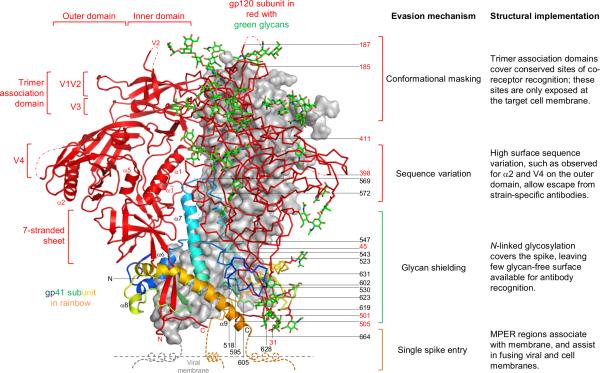
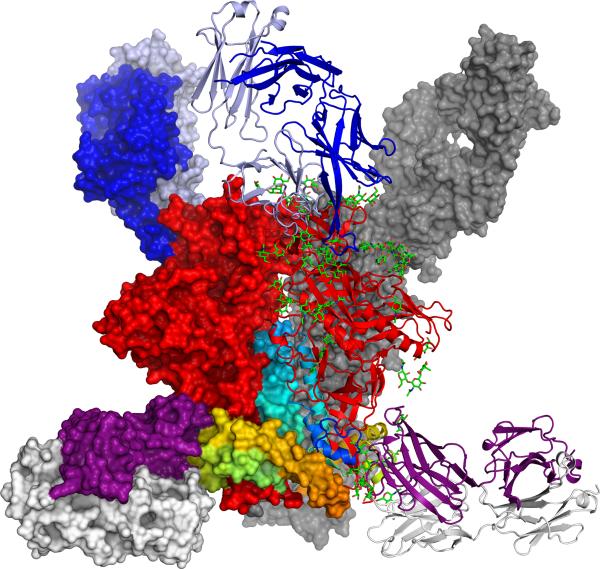
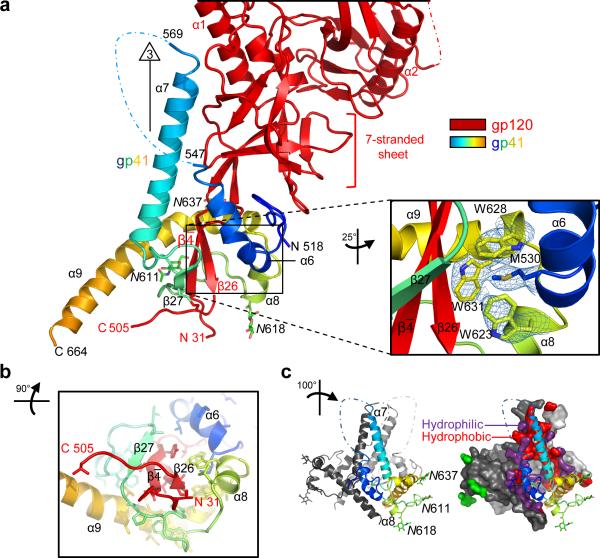
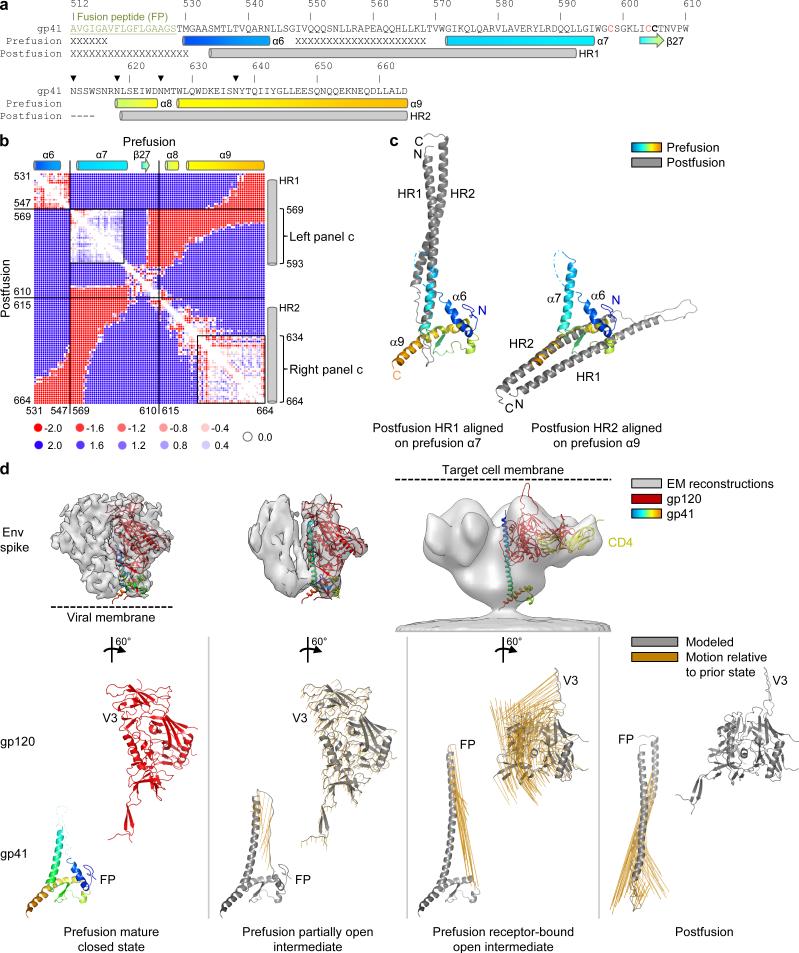
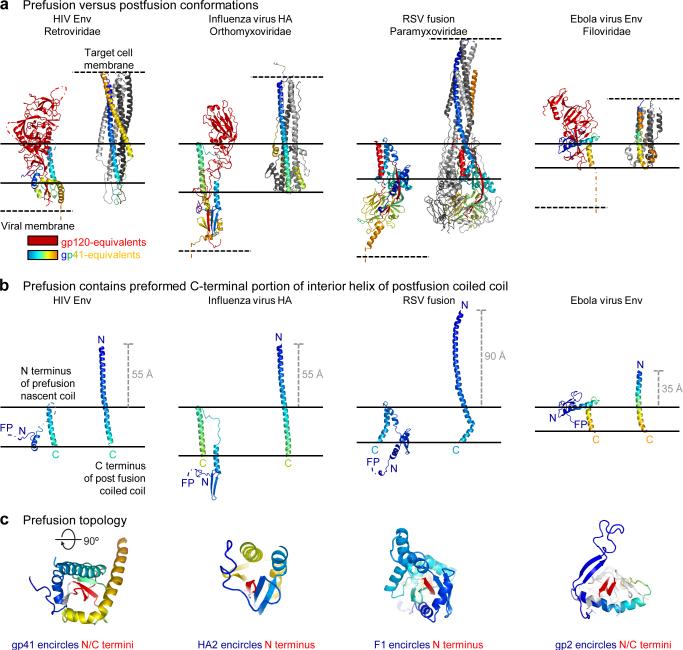
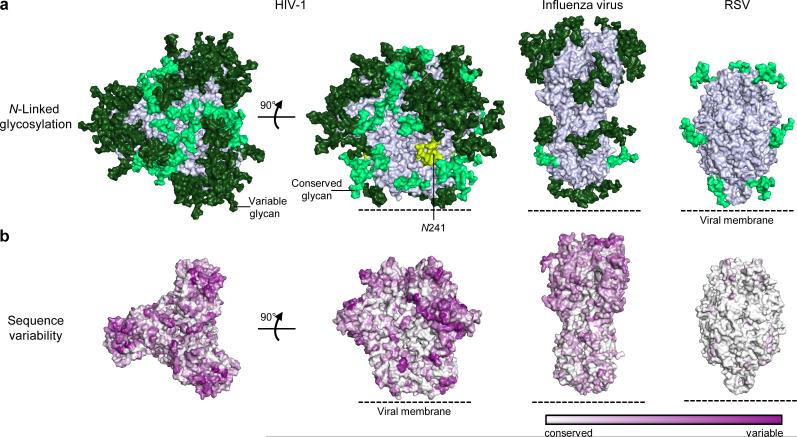
The human immunodeficiency virus type 1 (HIV-1) envelope (Env) spike, comprising three gp120 and three gp41 subunits, is a conformational machine that facilitates HIV-1 entry by rearranging from a mature unliganded state, through receptor-bound intermediates, to a post-fusion state. As the sole viral antigen on the HIV-1 virion surface, Env is both the target of neutralizing antibodies and a focus of vaccine efforts. Here we report the structure at 3.5 Å resolution for an HIV-1 Env trimer captured in a mature closed state by antibodies PGT122 and 35O22. This structure reveals the pre-fusion conformation of gp41, indicates rearrangements needed for fusion activation, and defines parameters of immune evasion and immune recognition. Pre-fusion gp41 encircles amino- and carboxy-terminal strands of gp120 with four helices that form a membrane-proximal collar, fastened by insertion of a fusion peptide-proximal methionine into a gp41-tryptophan clasp. Spike rearrangements required for entry involve opening the clasp and expelling the termini. N-linked glycosylation and sequence-variable regions cover the pre-fusion closed spike; we used chronic cohorts to map the prevalence and location of effective HIV-1-neutralizing responses, which were distinguished by their recognition of N-linked glycan and tolerance for epitope-sequence variation.
Figures



Comment in
-
HIV: A stamp on the envelope.Nature. 2014 Oct 23;514(7523):437-8. doi: 10.1038/nature13926. Epub 2014 Oct 8. Nature. 2014. PMID: 25296251 Free PMC article.
Similar articles
-
Structure and Recognition of a Novel HIV-1 gp120-gp41 Interface Antibody that Caused MPER Exposure through Viral Escape.PLoS Pathog. 2017 Jan 11;13(1):e1006074. doi: 10.1371/journal.ppat.1006074. eCollection 2017 Jan. PLoS Pathog. 2017. PMID: 28076415 Free PMC article.
-
Allosteric modulation of the HIV-1 gp120-gp41 association site by adjacent gp120 variable region 1 (V1) N-glycans linked to neutralization sensitivity.PLoS Pathog. 2013;9(4):e1003218. doi: 10.1371/journal.ppat.1003218. Epub 2013 Apr 4. PLoS Pathog. 2013. PMID: 23592978 Free PMC article.
-
Glycosylation Benchmark Profile for HIV-1 Envelope Glycoprotein Production Based on Eleven Env Trimers.J Virol. 2017 Apr 13;91(9):e02428-16. doi: 10.1128/JVI.02428-16. Print 2017 May 1. J Virol. 2017. PMID: 28202756 Free PMC article.
-
Aiming to induce broadly reactive neutralizing antibody responses with HIV-1 vaccine candidates.Expert Rev Vaccines. 2006 Jun;5(3):347-63. doi: 10.1586/14760584.5.3.347. Expert Rev Vaccines. 2006. PMID: 16827619 Free PMC article. Review.
-
Quaternary Interaction of the HIV-1 Envelope Trimer with CD4 and Neutralizing Antibodies.Viruses. 2021 Jul 20;13(7):1405. doi: 10.3390/v13071405. Viruses. 2021. PMID: 34372611 Free PMC article. Review.
Cited by
-
50 Years of structural immunology.J Biol Chem. 2021 Jan-Jun;296:100745. doi: 10.1016/j.jbc.2021.100745. Epub 2021 May 3. J Biol Chem. 2021. PMID: 33957119 Free PMC article. Review.
-
South African HIV-1 subtype C transmitted variants with a specific V2 motif show higher dependence on α4β7 for replication.Retrovirology. 2015 Jun 24;12:54. doi: 10.1186/s12977-015-0183-3. Retrovirology. 2015. PMID: 26105197 Free PMC article.
-
A Prominent Site of Antibody Vulnerability on HIV Envelope Incorporates a Motif Associated with CCR5 Binding and Its Camouflaging Glycans.Immunity. 2016 Jul 19;45(1):31-45. doi: 10.1016/j.immuni.2016.06.026. Immunity. 2016. PMID: 27438765 Free PMC article.
-
HIV-1 Envelope Glycosylation and the Signal Peptide.Vaccines (Basel). 2021 Feb 19;9(2):176. doi: 10.3390/vaccines9020176. Vaccines (Basel). 2021. PMID: 33669676 Free PMC article. Review.
-
Closing coronavirus spike glycoproteins by structure-guided design.bioRxiv [Preprint]. 2020 Jun 3:2020.06.03.129817. doi: 10.1101/2020.06.03.129817. bioRxiv. 2020. Update in: Nat Struct Mol Biol. 2020 Oct;27(10):942-949. doi: 10.1038/s41594-020-0483-8 PMID: 32577661 Free PMC article. Updated. Preprint.
References
-
- The Joint United Nations Programme on HIV/AIDS 2013 Report on the global AIDS epidemic. 2013
-
- Wyatt R, Sodroski J. The HIV-1 envelope glycoproteins: fusogens, antigens, and immunogens. Science. 1998;280:1884–1888. - PubMed
-
- Wei X, et al. Antibody neutralization and escape by HIV-1. Nature. 2003;422:307–312. - PubMed
-
- Leonard CK, et al. Assignment of intrachain disulfide bonds and characterization of potential glycosylation sites of the type 1 recombinant human immunodeficiency virus envelope glycoprotein (gp120) expressed in Chinese hamster ovary cells. The Journal of biological chemistry. 1990;265:10373–10382. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
- UM1 AI100645/AI/NIAID NIH HHS/United States
- R01 GM098859/GM/NIGMS NIH HHS/United States
- ZIA AI005023-13/ImNIH/Intramural NIH HHS/United States
- AI100645/AI/NIAID NIH HHS/United States
- P01 GM056550/GM/NIGMS NIH HHS/United States
- ZIA AI005024-13/ImNIH/Intramural NIH HHS/United States
- R21 AI100696/AI/NIAID NIH HHS/United States
- UL1 TR000142/TR/NCATS NIH HHS/United States
- AI0678501/AI/NIAID NIH HHS/United States
- P30 AI050410/AI/NIAID NIH HHS/United States
- R01-GM098859/GM/NIGMS NIH HHS/United States
- P30 CA016086/CA/NCI NIH HHS/United States
- P01-GM56550/GM/NIGMS NIH HHS/United States
- R21-AI100696/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
