

Extracellular matrix regulation of inflammation in the healthy and injured spinal cord
- PMID: 25017885
- PMCID: PMC4099942
- DOI: 10.1016/j.expneurol.2013.11.020
Extracellular matrix regulation of inflammation in the healthy and injured spinal cord
Abstract
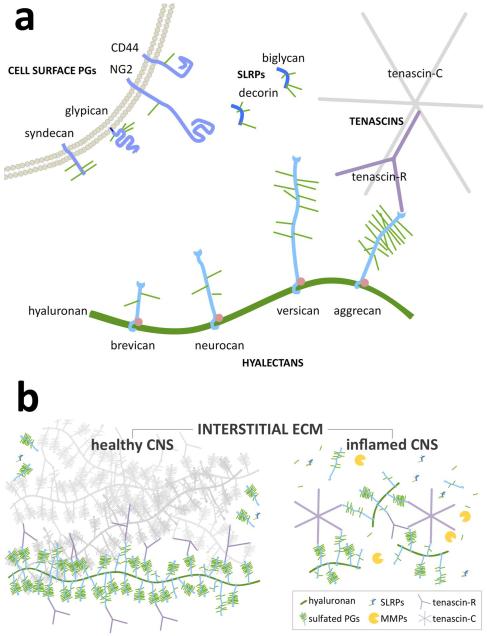
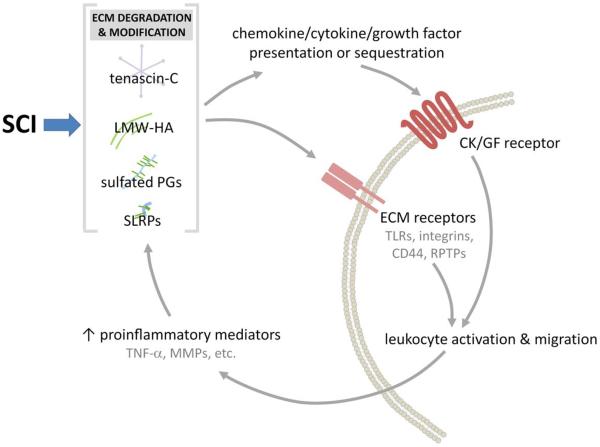
Throughout the body, the extracellular matrix (ECM) provides structure and organization to tissues and also helps regulate cell migration and intercellular communication. In the injured spinal cord (or brain), changes in the composition and structure of the ECM undoubtedly contribute to regeneration failure. Less appreciated is how the native and injured ECM influences intraspinal inflammation and, conversely, how neuroinflammation affects the synthesis and deposition of ECM after CNS injury. In all tissues, inflammation can be initiated and propagated by ECM disruption. Molecules of ECM newly liberated by injury or inflammation include hyaluronan fragments, tenascins, and sulfated proteoglycans. These act as "damage-associated molecular patterns" or "alarmins", i.e., endogenous proteins that trigger and subsequently amplify inflammation. Activated inflammatory cells, in turn, further damage the ECM by releasing degradative enzymes including matrix metalloproteinases (MMPs). After spinal cord injury (SCI), destabilization or alteration of the structural and chemical compositions of the ECM affects migration, communication, and survival of all cells - neural and non-neural - that are critical for spinal cord repair. By stabilizing ECM structure or modifying their ability to trigger the degradative effects of inflammation, it may be possible to create an environment that is more conducive to tissue repair and axon plasticity after SCI.
Keywords: DAMP; Hyaluronan; Immune; Neuroinflammation; Proteoglycan; TLR; Tenascin; Toll-like receptors.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures

Similar articles
-
miR-155 Deletion in Mice Overcomes Neuron-Intrinsic and Neuron-Extrinsic Barriers to Spinal Cord Repair.J Neurosci. 2016 Aug 10;36(32):8516-32. doi: 10.1523/JNEUROSCI.0735-16.2016. J Neurosci. 2016. PMID: 27511021 Free PMC article.
-
Harnessing developmental dynamics of spinal cord extracellular matrix improves regenerative potential of spinal cord organoids.Cell Stem Cell. 2024 May 2;31(5):772-787.e11. doi: 10.1016/j.stem.2024.03.007. Epub 2024 Apr 1. Cell Stem Cell. 2024. PMID: 38565140
-
Extracellular Matrix and Oxidative Stress Following Traumatic Spinal Cord Injury: Physiological and Pathophysiological Roles and Opportunities for Therapeutic Intervention.Antioxid Redox Signal. 2022 Jul;37(1-3):184-207. doi: 10.1089/ars.2021.0120. Epub 2021 Dec 7. Antioxid Redox Signal. 2022. PMID: 34465134 Review.
-
Manipulating the extracellular matrix and its role in brain and spinal cord plasticity and repair.Neuropathol Appl Neurobiol. 2014 Feb;40(1):26-59. doi: 10.1111/nan.12114. Neuropathol Appl Neurobiol. 2014. PMID: 24438519 Review.
-
Neutrophil, Extracellular Matrix Components, and Their Interlinked Action in Promoting Secondary Pathogenesis After Spinal Cord Injury.Mol Neurobiol. 2021 Sep;58(9):4652-4665. doi: 10.1007/s12035-021-02443-5. Epub 2021 Jun 22. Mol Neurobiol. 2021. PMID: 34159551 Review.
Cited by
-
Curcumin, inflammation, and chronic diseases: how are they linked?Molecules. 2015 May 20;20(5):9183-213. doi: 10.3390/molecules20059183. Molecules. 2015. PMID: 26007179 Free PMC article. Review.
-
Epigenetic regulation and factors that influence the effect of iPSCs-derived neural stem/progenitor cells (NS/PCs) in the treatment of spinal cord injury.Clin Epigenetics. 2024 Feb 21;16(1):30. doi: 10.1186/s13148-024-01639-5. Clin Epigenetics. 2024. PMID: 38383473 Free PMC article. Review.
-
Decellularization alters the unfavorable regenerative adverse microenvironment of the injured spinal cord to support neurite outgrowth.Ann Transl Med. 2022 Sep;10(17):934. doi: 10.21037/atm-22-3969. Ann Transl Med. 2022. PMID: 36172103 Free PMC article.
-
Extracellular matrix composition determines astrocyte responses to mechanical and inflammatory stimuli.Neurosci Lett. 2015 Jul 23;600:104-9. doi: 10.1016/j.neulet.2015.06.013. Epub 2015 Jun 9. Neurosci Lett. 2015. PMID: 26067407 Free PMC article.
-
Interaction between integrin α5 and PDE4D regulates endothelial inflammatory signalling.Nat Cell Biol. 2016 Oct;18(10):1043-53. doi: 10.1038/ncb3405. Epub 2016 Sep 5. Nat Cell Biol. 2016. PMID: 27595237 Free PMC article.
References
-
- Agrawal SM, Lau L, Yong VW. MMPs in the central nervous system: where the good guys go bad. Semin. Cell Dev. Biol. 2008;19:42–51. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
