

Prolactin (PRL)-stimulated ubiquitination of ZnT2 mediates a transient increase in zinc secretion followed by ZnT2 degradation in mammary epithelial cells
- PMID: 25016022
- PMCID: PMC4156067
- DOI: 10.1074/jbc.M113.531145
Prolactin (PRL)-stimulated ubiquitination of ZnT2 mediates a transient increase in zinc secretion followed by ZnT2 degradation in mammary epithelial cells
Abstract
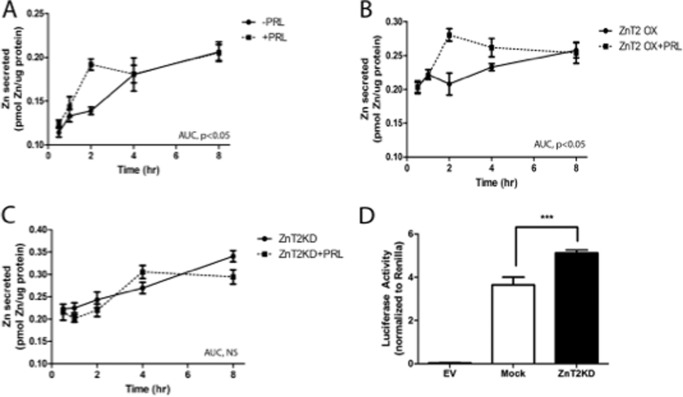
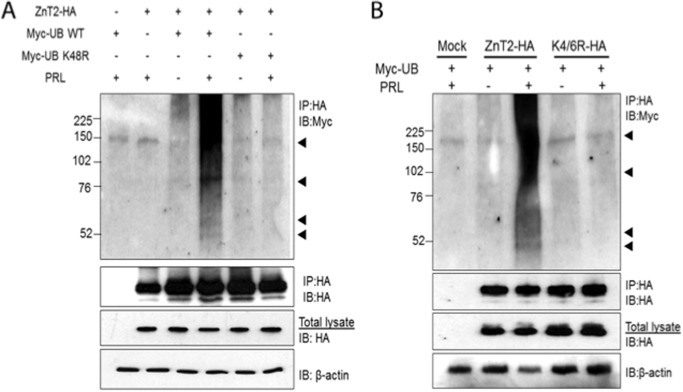
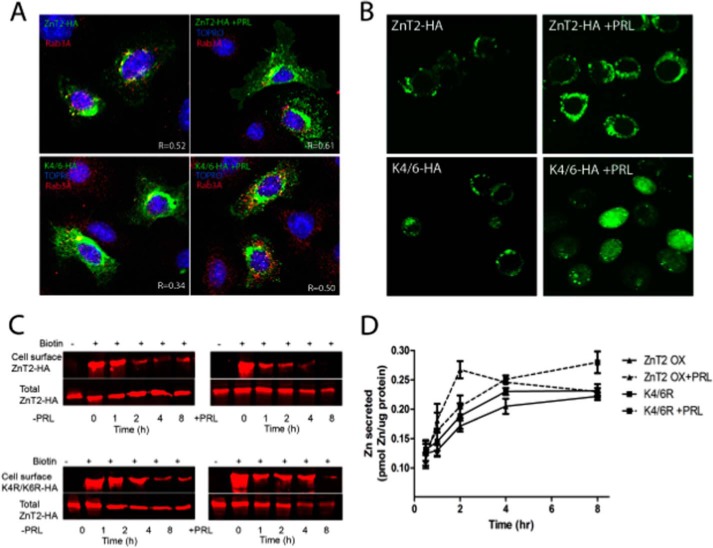
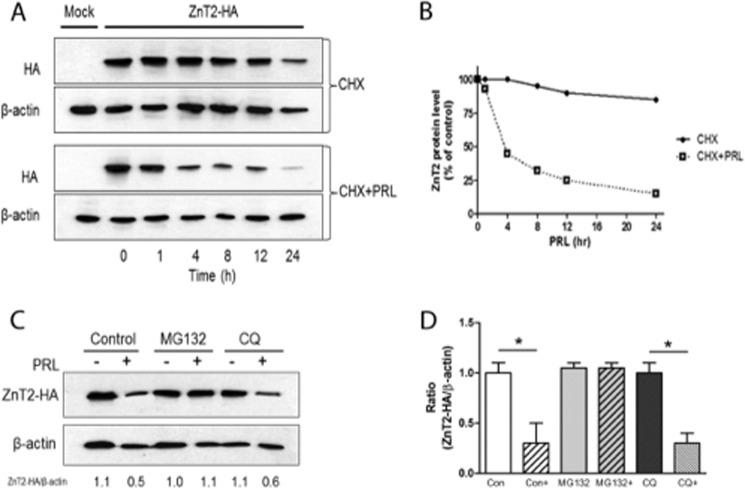
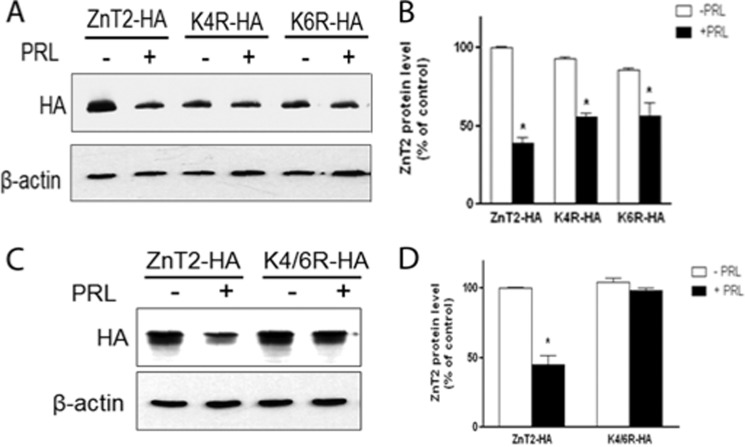
The zinc transporter ZnT2 imports zinc into secretory vesicles and regulates zinc export from the mammary epithelial cell. Mutations in ZnT2 substantially impair zinc secretion into milk. The lactogenic hormone prolactin (PRL) transcriptionally increases ZnT2 expression through the Jak2/STAT5 signaling pathway, increasing zinc accumulation in secretory vesicles and zinc secretion. Herein, we report that PRL post-translationally stimulated ZnT2 ubiquitination, which altered ZnT2 trafficking and augmented vesicular zinc accumulation and secretion from mammary epithelial cells in a transient manner. Ubiquitination then down-regulated zinc secretion by stimulating degradation of ZnT2. Mutagenesis of two N-terminal lysine residues (K4R and K6R) inhibited ZnT2 ubiquitination, vesicular zinc accumulation and secretion, and protein degradation. These findings establish that PRL post-translationally regulates ZnT2-mediated zinc secretion in a multifactorial manner, first by enhancing zinc accumulation in vesicles to transiently enhance zinc secretion and then by activating ubiquitin-dependent ZnT2 degradation. This provides insight into novel mechanisms through which ZnT2 and zinc transport is tightly regulated in mammary epithelial cells.
Keywords: Mammary Gland; Prolactin; Protein Degradation; Secretion; Ubiquitination; Zinc Transporter; ZnT2.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
Similar articles
-
Essential Role for Zinc Transporter 2 (ZnT2)-mediated Zinc Transport in Mammary Gland Development and Function during Lactation.J Biol Chem. 2015 May 22;290(21):13064-78. doi: 10.1074/jbc.M115.637439. Epub 2015 Apr 7. J Biol Chem. 2015. PMID: 25851903 Free PMC article.
-
Prolactin regulates ZNT2 expression through the JAK2/STAT5 signaling pathway in mammary cells.Am J Physiol Cell Physiol. 2009 Aug;297(2):C369-77. doi: 10.1152/ajpcell.00589.2008. Epub 2009 Jun 3. Am J Physiol Cell Physiol. 2009. PMID: 19494234 Free PMC article.
-
Zinc transporter 2 interacts with vacuolar ATPase and is required for polarization, vesicle acidification, and secretion in mammary epithelial cells.J Biol Chem. 2017 Dec 29;292(52):21598-21613. doi: 10.1074/jbc.M117.794461. Epub 2017 Nov 7. J Biol Chem. 2017. PMID: 29114036 Free PMC article.
-
New insights into the importance of prolactin in dairy ruminants.J Dairy Sci. 2016 Jan;99(1):864-74. doi: 10.3168/jds.2015-10035. Epub 2015 Nov 5. J Dairy Sci. 2016. PMID: 26547648 Review.
-
Prolactin and mammary gland development.J Mammary Gland Biol Neoplasia. 1999 Jan;4(1):79-88. doi: 10.1023/a:1018708704335. J Mammary Gland Biol Neoplasia. 1999. PMID: 10219908 Review.
Cited by
-
Role of zinc in health and disease.Clin Exp Med. 2024 Feb 17;24(1):38. doi: 10.1007/s10238-024-01302-6. Clin Exp Med. 2024. PMID: 38367035 Free PMC article. Review.
-
Zinc-sensitive MRI contrast agent detects differential release of Zn(II) ions from the healthy vs. malignant mouse prostate.Proc Natl Acad Sci U S A. 2016 Sep 13;113(37):E5464-71. doi: 10.1073/pnas.1609450113. Epub 2016 Aug 25. Proc Natl Acad Sci U S A. 2016. PMID: 27562169 Free PMC article.
-
Zinc transporters and their functional integration in mammalian cells.J Biol Chem. 2021 Jan-Jun;296:100320. doi: 10.1016/j.jbc.2021.100320. Epub 2021 Jan 22. J Biol Chem. 2021. PMID: 33485965 Free PMC article. Review.
-
Essential Role for Zinc Transporter 2 (ZnT2)-mediated Zinc Transport in Mammary Gland Development and Function during Lactation.J Biol Chem. 2015 May 22;290(21):13064-78. doi: 10.1074/jbc.M115.637439. Epub 2015 Apr 7. J Biol Chem. 2015. PMID: 25851903 Free PMC article.
-
A genetic variant in SLC30A2 causes breast dysfunction during lactation by inducing ER stress, oxidative stress and epithelial barrier defects.Sci Rep. 2018 Feb 23;8(1):3542. doi: 10.1038/s41598-018-21505-8. Sci Rep. 2018. PMID: 29476070 Free PMC article.
References
-
- Chowanadisai W., Lönnerdal B., Kelleher S. L. (2006) Identification of a mutation in SLC30A2 (ZnT-2) in women with low milk zinc concentration that results in transient neonatal zinc deficiency. J. Biol. Chem. 281, 39699–39707 - PubMed
-
- Lasry I., Seo Y. A., Ityel H., Shalva N., Pode-Shakked B., Glaser F., Berman B., Berezovsky I., Goncearenco A., Klar A., Levy J., Anikster Y., Kelleher S. L., Assaraf Y. G. (2012) A dominant negative heterozygous G87R mutation in the zinc transporter, ZnT-2 (SLC30A2), results in transient neonatal zinc deficiency. J. Biol. Chem. 287, 29348–29361 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
