

Toxin instability and its role in toxin translocation from the endoplasmic reticulum to the cytosol
- PMID: 24970201
- PMCID: PMC4030972
- DOI: 10.3390/biom3040997
Toxin instability and its role in toxin translocation from the endoplasmic reticulum to the cytosol
Abstract
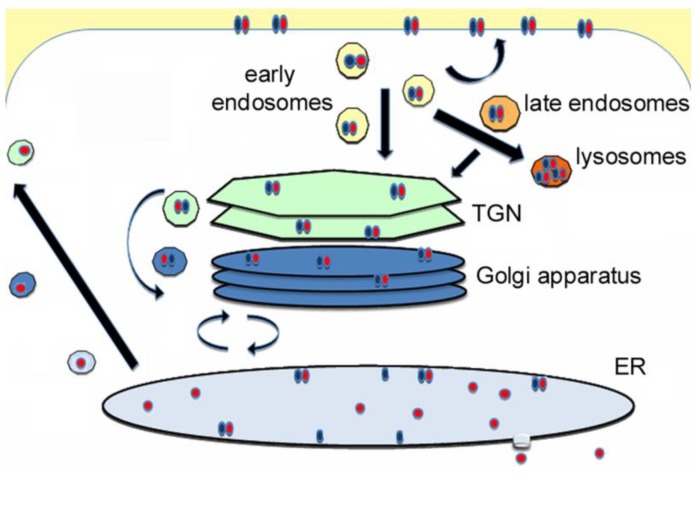
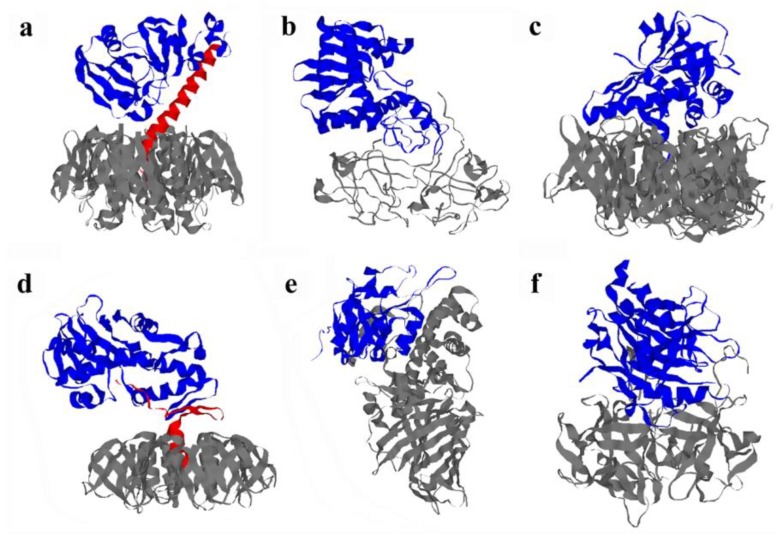
AB toxins enter a host cell by receptor-mediated endocytosis. The catalytic A chain then crosses the endosome or endoplasmic reticulum (ER) membrane to reach its cytosolic target. Dissociation of the A chain from the cell-binding B chain occurs before or during translocation to the cytosol, and only the A chain enters the cytosol. In some cases, AB subunit dissociation is facilitated by the unique physiology and function of the ER. The A chains of these ER-translocating toxins are stable within the architecture of the AB holotoxin, but toxin disassembly results in spontaneous or assisted unfolding of the isolated A chain. This unfolding event places the A chain in a translocation-competent conformation that promotes its export to the cytosol through the quality control mechanism of ER-associated degradation. A lack of lysine residues for ubiquitin conjugation protects the exported A chain from degradation by the ubiquitin-proteasome system, and an interaction with host factors allows the cytosolic toxin to regain a folded, active state. The intrinsic instability of the toxin A chain thus influences multiple steps of the intoxication process. This review will focus on the host-toxin interactions involved with A chain unfolding in the ER and A chain refolding in the cytosol.
Figures
Similar articles
-
Thermal Unfolding of the Pertussis Toxin S1 Subunit Facilitates Toxin Translocation to the Cytosol by the Mechanism of Endoplasmic Reticulum-Associated Degradation.Infect Immun. 2016 Nov 18;84(12):3388-3398. doi: 10.1128/IAI.00732-16. Print 2016 Dec. Infect Immun. 2016. PMID: 27647866 Free PMC article.
-
A binding motif for Hsp90 in the A chains of ADP-ribosylating toxins that move from the endoplasmic reticulum to the cytosol.Cell Microbiol. 2019 Oct;21(10):e13074. doi: 10.1111/cmi.13074. Epub 2019 Jul 5. Cell Microbiol. 2019. PMID: 31231933 Free PMC article.
-
The low lysine content of ricin A chain reduces the risk of proteolytic degradation after translocation from the endoplasmic reticulum to the cytosol.Biochemistry. 2002 Mar 12;41(10):3405-13. doi: 10.1021/bi011580v. Biochemistry. 2002. PMID: 11876649
-
Intracellular Trafficking and Translocation of Pertussis Toxin.Toxins (Basel). 2019 Jul 25;11(8):437. doi: 10.3390/toxins11080437. Toxins (Basel). 2019. PMID: 31349590 Free PMC article. Review.
-
How ricin and Shiga toxin reach the cytosol of target cells: retrotranslocation from the endoplasmic reticulum.Curr Top Microbiol Immunol. 2012;357:19-40. doi: 10.1007/82_2011_154. Curr Top Microbiol Immunol. 2012. PMID: 21761287 Review.
Cited by
-
A Conformational Shift in the Dissociated Cholera Toxin A1 Subunit Prevents Reassembly of the Cholera Holotoxin.Toxins (Basel). 2015 Jul 20;7(7):2674-84. doi: 10.3390/toxins7072674. Toxins (Basel). 2015. PMID: 26266549 Free PMC article.
-
Differences in Medium-Induced Conformational Plasticity Presumably Underlie Different Cytotoxic Activity of Ricin and Viscumin.Biomolecules. 2022 Feb 11;12(2):295. doi: 10.3390/biom12020295. Biomolecules. 2022. PMID: 35204796 Free PMC article.
-
Cellular recovery from exposure to sub-optimal concentrations of AB toxins that inhibit protein synthesis.Sci Rep. 2018 Feb 6;8(1):2494. doi: 10.1038/s41598-018-20861-9. Sci Rep. 2018. PMID: 29410492 Free PMC article.
-
Toxins Utilize the Endoplasmic Reticulum-Associated Protein Degradation Pathway in Their Intoxication Process.Int J Mol Sci. 2019 Mar 15;20(6):1307. doi: 10.3390/ijms20061307. Int J Mol Sci. 2019. PMID: 30875878 Free PMC article. Review.
-
Thermal Unfolding of the Pertussis Toxin S1 Subunit Facilitates Toxin Translocation to the Cytosol by the Mechanism of Endoplasmic Reticulum-Associated Degradation.Infect Immun. 2016 Nov 18;84(12):3388-3398. doi: 10.1128/IAI.00732-16. Print 2016 Dec. Infect Immun. 2016. PMID: 27647866 Free PMC article.
References
-
- Barth H. Exploring the role of host cell chaperones/PPIases during cellular up-take of bacterial ADP-ribosylating toxins as basis for novel pharmacological strategies to protect mammalian cells against these virulence factors. Naunyn. Schmiedebergs. Arch. Pharmacol. 2011;383:237–245. doi: 10.1007/s00210-010-0581-y. - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
