

The Mammalian response to virus infection is independent of small RNA silencing
- PMID: 24953656
- PMCID: PMC4096324
- DOI: 10.1016/j.celrep.2014.05.038
The Mammalian response to virus infection is independent of small RNA silencing
Abstract
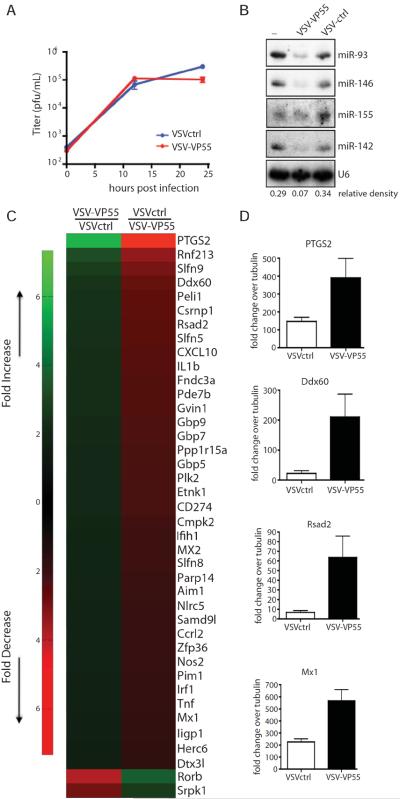
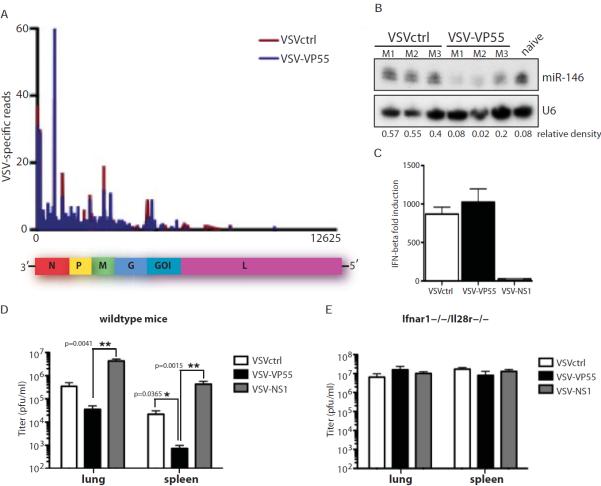
A successful cellular response to virus infection is essential for evolutionary survival. In plants, arthropods, and nematodes, cellular antiviral defenses rely on RNAi. Interestingly, the mammalian response to virus is predominantly orchestrated through interferon (IFN)-mediated induction of antiviral proteins. Despite the potency of the IFN system, it remains unclear whether mammals also have the capacity to employ antiviral RNAi. Here, we investigated this by disabling IFN function, small RNA function, or both activities in the context of virus infection. We find that loss of small RNAs in the context of an in vivo RNA virus infection lowers titers due to reduced transcriptional repression of the host antiviral response. In contrast, enabling a virus with the capacity to inhibit the IFN system results in increased titers. Taken together, these results indicate that small RNA silencing is not a physiological contributor to the IFN-mediated cellular response to virus infection.
Copyright © 2014 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
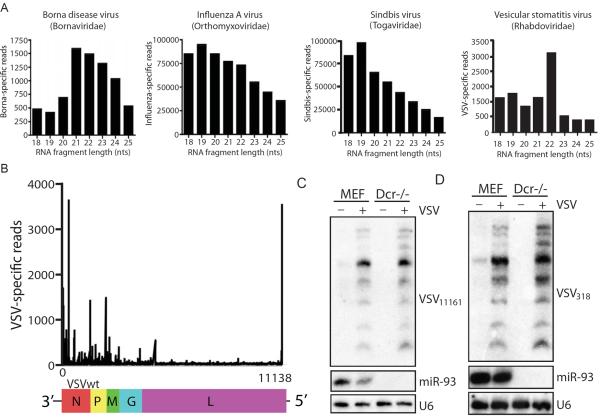
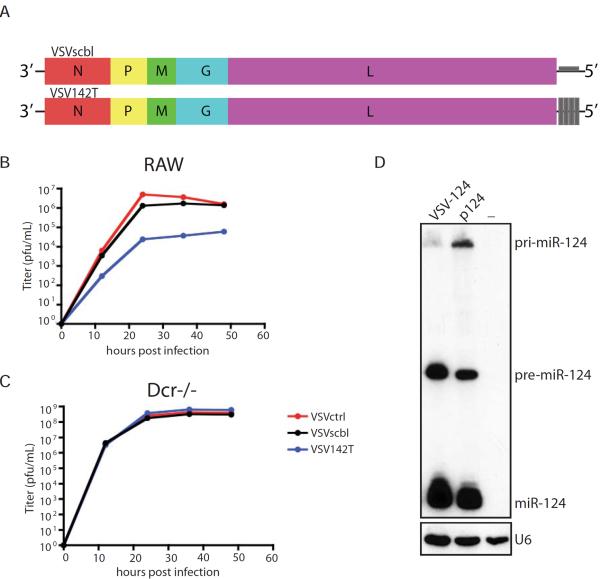
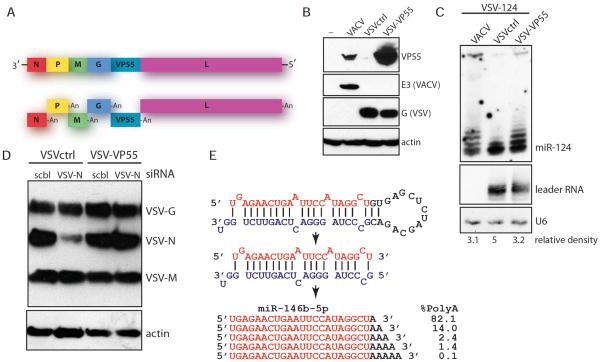
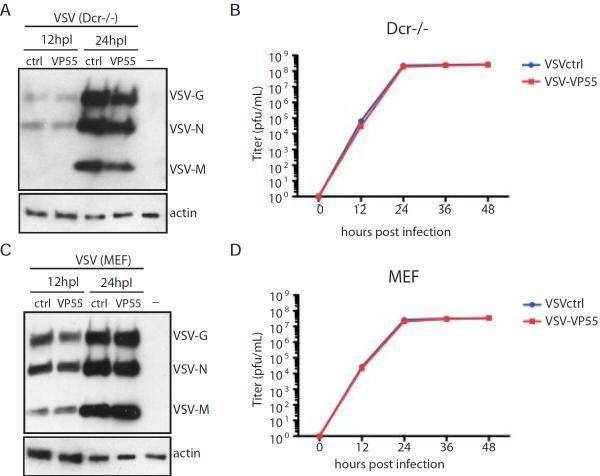
Similar articles
-
Chandipura Virus Utilizes the Prosurvival Function of RelA NF-κB for Its Propagation.J Virol. 2019 Jun 28;93(14):e00081-19. doi: 10.1128/JVI.00081-19. Print 2019 Jul 15. J Virol. 2019. PMID: 31043529 Free PMC article.
-
Caenorhabditis elegans RIG-I Homolog Mediates Antiviral RNA Interference Downstream of Dicer-Dependent Biogenesis of Viral Small Interfering RNAs.mBio. 2017 Mar 21;8(2):e00264-17. doi: 10.1128/mBio.00264-17. mBio. 2017. PMID: 28325765 Free PMC article.
-
Antiviral innate immune response of RNA interference.J Infect Dev Ctries. 2014 Jul 14;8(7):804-10. doi: 10.3855/jidc.4187. J Infect Dev Ctries. 2014. PMID: 25022288 Review.
-
Mutual Regulation of RNA Silencing and the IFN Response as an Antiviral Defense System in Mammalian Cells.Int J Mol Sci. 2020 Feb 17;21(4):1348. doi: 10.3390/ijms21041348. Int J Mol Sci. 2020. PMID: 32079277 Free PMC article. Review.
-
Dicer is involved in protection against influenza A virus infection.J Gen Virol. 2007 Oct;88(Pt 10):2627-2635. doi: 10.1099/vir.0.83103-0. J Gen Virol. 2007. PMID: 17872512
Cited by
-
Toll-like receptor-3 is dispensable for the innate microRNA response to West Nile virus (WNV).PLoS One. 2014 Aug 15;9(8):e104770. doi: 10.1371/journal.pone.0104770. eCollection 2014. PLoS One. 2014. PMID: 25127040 Free PMC article.
-
Cross-species comparative analysis of Dicer proteins during Sindbis virus infection.Sci Rep. 2015 May 29;5:10693. doi: 10.1038/srep10693. Sci Rep. 2015. PMID: 26024431 Free PMC article.
-
Prediction and Analysis of SARS-CoV-2-Targeting MicroRNA in Human Lung Epithelium.Genes (Basel). 2020 Aug 26;11(9):1002. doi: 10.3390/genes11091002. Genes (Basel). 2020. PMID: 32858958 Free PMC article.
-
Antiviral RNA interference in mammals.Curr Opin Immunol. 2018 Oct;54:109-114. doi: 10.1016/j.coi.2018.06.010. Epub 2018 Jul 17. Curr Opin Immunol. 2018. PMID: 30015086 Free PMC article. Review.
-
STUB1 regulates antiviral RNAi through inducing ubiquitination and degradation of Dicer and AGO2 in mammals.Virol Sin. 2022 Aug;37(4):569-580. doi: 10.1016/j.virs.2022.05.001. Epub 2022 May 6. Virol Sin. 2022. PMID: 35533808 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
