

PD-L1 is a novel direct target of HIF-1α, and its blockade under hypoxia enhanced MDSC-mediated T cell activation
- PMID: 24778419
- PMCID: PMC4010891
- DOI: 10.1084/jem.20131916
PD-L1 is a novel direct target of HIF-1α, and its blockade under hypoxia enhanced MDSC-mediated T cell activation
Abstract
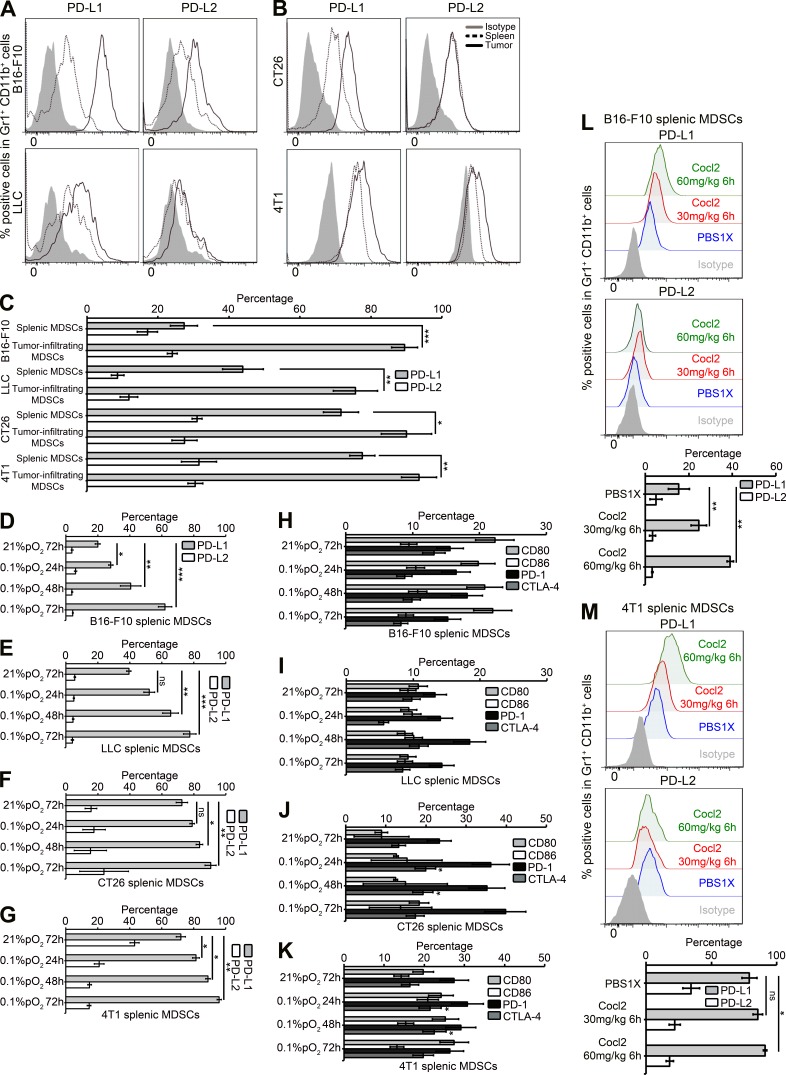
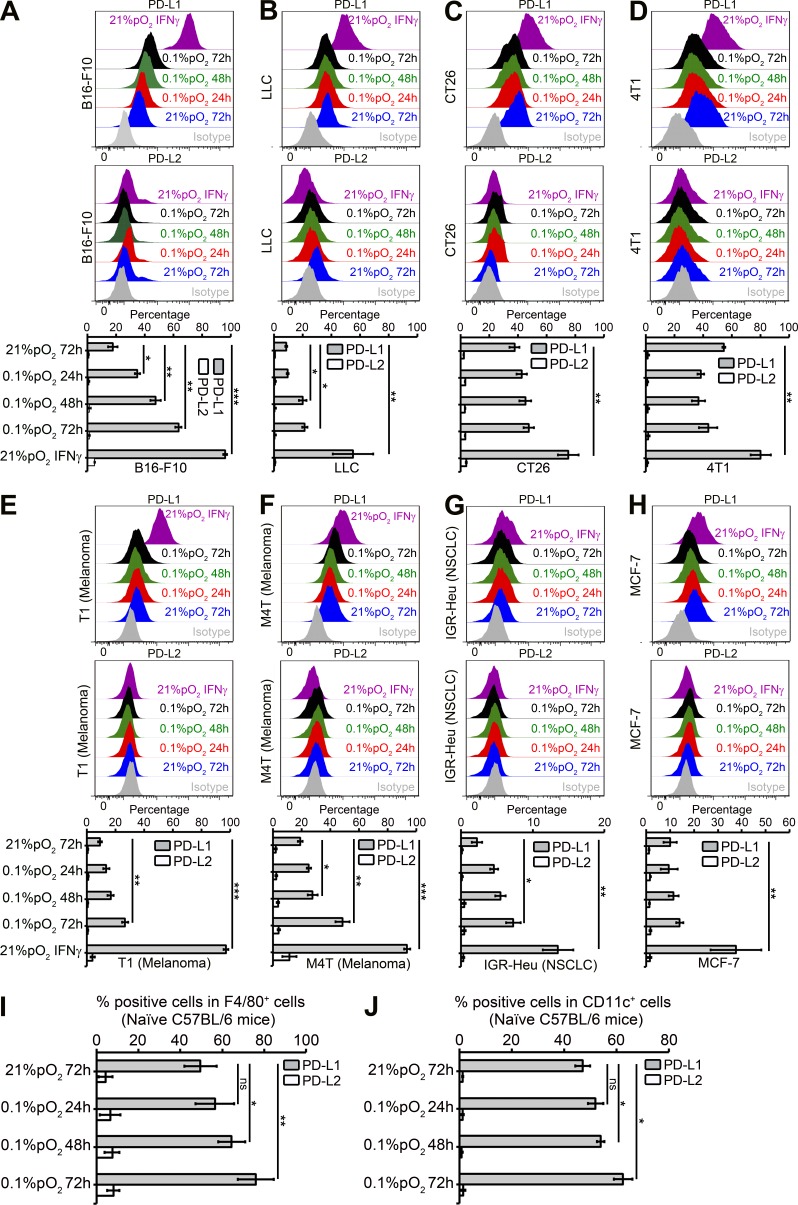
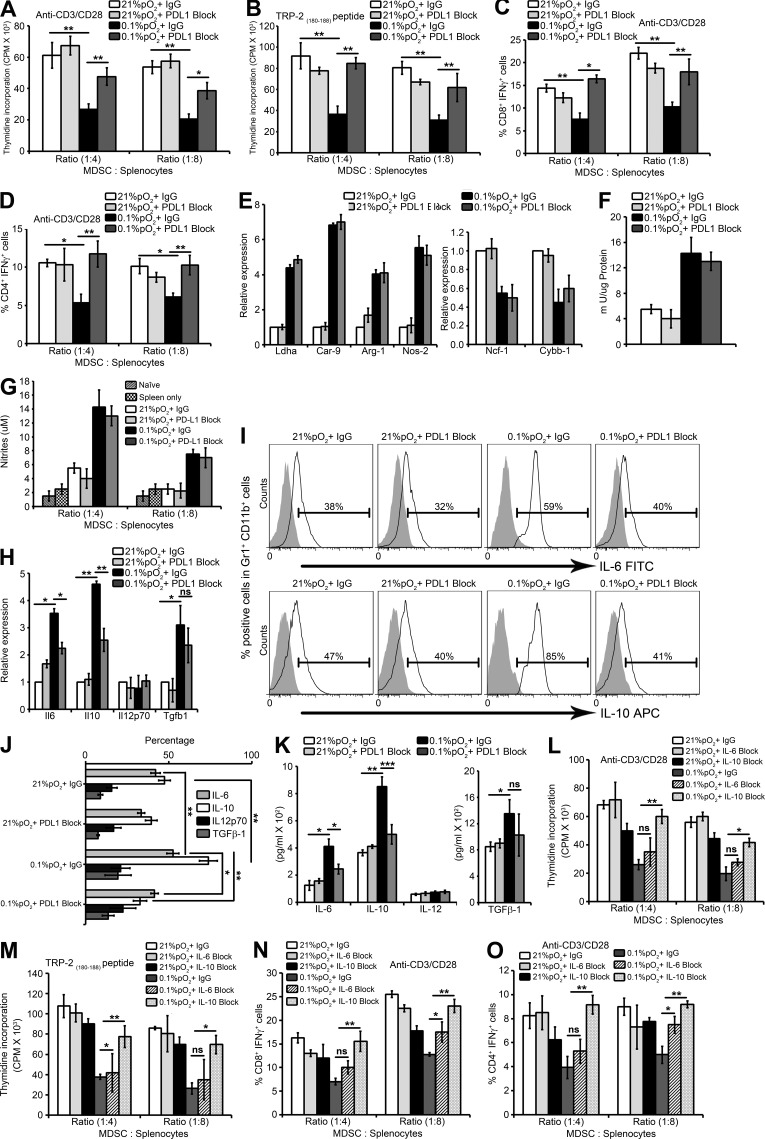
Tumor-infiltrating myeloid cells such as myeloid-derived suppressor cells (MDSCs) and tumor-associated macrophages (TAMs) form an important component of the hypoxic tumor microenvironment. Here, we investigated the influence of hypoxia on immune checkpoint receptors (programmed death [PD]-1 and CTLA-4) and their respective ligands (PD-1 ligand 1 [PD-L1], PD-L2, CD80, and CD86) on MDSCs. We demonstrate that MDSCs at the tumor site show a differential expression of PD-L1 as compared with MDSCs from peripheral lymphoid organ (spleen). Hypoxia caused a rapid, dramatic, and selective up-regulation of PD-L1 on splenic MDSCs in tumor-bearing mice. This was not limited to MDSCs, as hypoxia also significantly increased the expression of PD-L1 on macrophages, dendritic cells, and tumor cells. Furthermore, PD-L1 up-regulation under hypoxia was dependent on hypoxia-inducible factor-1α (HIF-1α) but not HIF-2α. Chromatin immunoprecipitation and luciferase reporter assay revealed direct binding of HIF-1α to a transcriptionally active hypoxia-response element (HRE) in the PD-L1 proximal promoter. Blockade of PD-L1 under hypoxia enhanced MDSC-mediated T cell activation and was accompanied by the down-regulation of MDSCs IL-6 and IL-10. Finally, neutralizing antibodies against IL-10 under hypoxia significantly abrogated the suppressive activity of MDSCs. Simultaneous blockade of PD-L1 along with inhibition of HIF-1α may thus represent a novel approach for cancer immunotherapy.
Figures

Similar articles
-
Decursin promotes HIF-1α proteasomal degradation and immune responses in hypoxic tumour microenvironment.Phytomedicine. 2020 Nov;78:153318. doi: 10.1016/j.phymed.2020.153318. Epub 2020 Sep 1. Phytomedicine. 2020. PMID: 32896707
-
Hypoxia-responsive lncRNA MIR155HG promotes PD-L1 expression in hepatocellular carcinoma cells by enhancing HIF-1α mRNA stability.Int Immunopharmacol. 2024 Jul 30;136:112415. doi: 10.1016/j.intimp.2024.112415. Epub 2024 Jun 7. Int Immunopharmacol. 2024. PMID: 38850791
-
Targeting hypoxia at the forefront of anticancer immune responses.Oncoimmunology. 2015 Jan 7;3(12):e954463. doi: 10.4161/21624011.2014.954463. eCollection 2014 Dec. Oncoimmunology. 2015. PMID: 25964858 Free PMC article.
-
Regulation of PD-L1: a novel role of pro-survival signalling in cancer.Ann Oncol. 2016 Mar;27(3):409-16. doi: 10.1093/annonc/mdv615. Epub 2015 Dec 17. Ann Oncol. 2016. PMID: 26681673 Review.
-
[Immunomodulation in the tumor microenvironment: Therapeutic potential of combined inhibition of tumor hypoxia and PD-1/ PD-L1].Magy Onkol. 2024 Jul 16;68(2):126-135. Epub 2024 Jun 3. Magy Onkol. 2024. PMID: 39013086 Review. Hungarian.
Cited by
-
Neutrophil diversity and plasticity in tumour progression and therapy.Nat Rev Cancer. 2020 Sep;20(9):485-503. doi: 10.1038/s41568-020-0281-y. Epub 2020 Jul 21. Nat Rev Cancer. 2020. PMID: 32694624 Review.
-
NK cells and solid tumors: therapeutic potential and persisting obstacles.Mol Cancer. 2022 Nov 1;21(1):206. doi: 10.1186/s12943-022-01672-z. Mol Cancer. 2022. PMID: 36319998 Free PMC article. Review.
-
Polarization and β-Glucan Reprogram Immunomodulatory Metabolism in Human Macrophages and Ex Vivo in Human Lung Cancer Tissues.J Immunol. 2022 Nov 1;209(9):1674-1690. doi: 10.4049/jimmunol.2200178. Epub 2022 Sep 23. J Immunol. 2022. PMID: 36150727 Free PMC article.
-
Continuous activation of polymorphonuclear myeloid-derived suppressor cells during pregnancy is critical for fetal development.Cell Mol Immunol. 2021 Jul;18(7):1692-1707. doi: 10.1038/s41423-021-00704-w. Epub 2021 Jun 7. Cell Mol Immunol. 2021. PMID: 34099889 Free PMC article.
-
Transcription factor HIF1A: downstream targets, associated pathways, polymorphic hypoxia response element (HRE) sites, and initiative for standardization of reporting in scientific literature.Tumour Biol. 2016 Nov;37(11):14851-14861. doi: 10.1007/s13277-016-5331-4. Epub 2016 Sep 19. Tumour Biol. 2016. PMID: 27644243
References
-
- Apolloni E., Bronte V., Mazzoni A., Serafini P., Cabrelle A., Segal D.M., Young H.A., Zanovello P. 2000. Immortalized myeloid suppressor cells trigger apoptosis in antigen-activated T lymphocytes. J. Immunol. 165:6723–6730 - PubMed
-
- Clambey E.T., McNamee E.N., Westrich J.A., Glover L.E., Campbell E.L., Jedlicka P., de Zoeten E.F., Cambier J.C., Stenmark K.R., Colgan S.P., Eltzschig H.K. 2012. Hypoxia-inducible factor-1 alpha-dependent induction of FoxP3 drives regulatory T-cell abundance and function during inflammatory hypoxia of the mucosa. Proc. Natl. Acad. Sci. USA. 109:E2784–E2793 10.1073/pnas.1202366109 - DOI - PMC - PubMed
-
- Corzo C.A., Condamine T., Lu L., Cotter M.J., Youn J.I., Cheng P., Cho H.I., Celis E., Quiceno D.G., Padhya T., et al. 2010. HIF-1α regulates function and differentiation of myeloid-derived suppressor cells in the tumor microenvironment. J. Exp. Med. 207:2439–2453 10.1084/jem.20100587 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
