

Emergence of the A20/ABIN-mediated inhibition of NF-κB signaling via modifying the ubiquitinated proteins in a basal chordate
- PMID: 24753567
- PMCID: PMC4020044
- DOI: 10.1073/pnas.1321187111
Emergence of the A20/ABIN-mediated inhibition of NF-κB signaling via modifying the ubiquitinated proteins in a basal chordate
Abstract
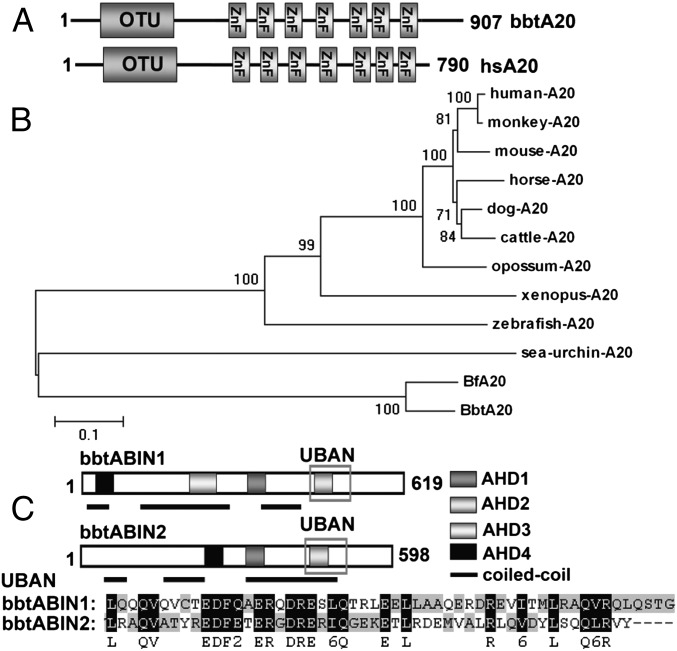
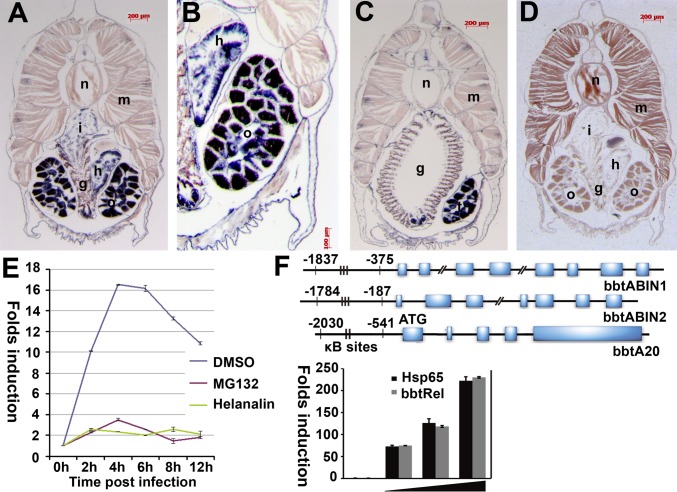
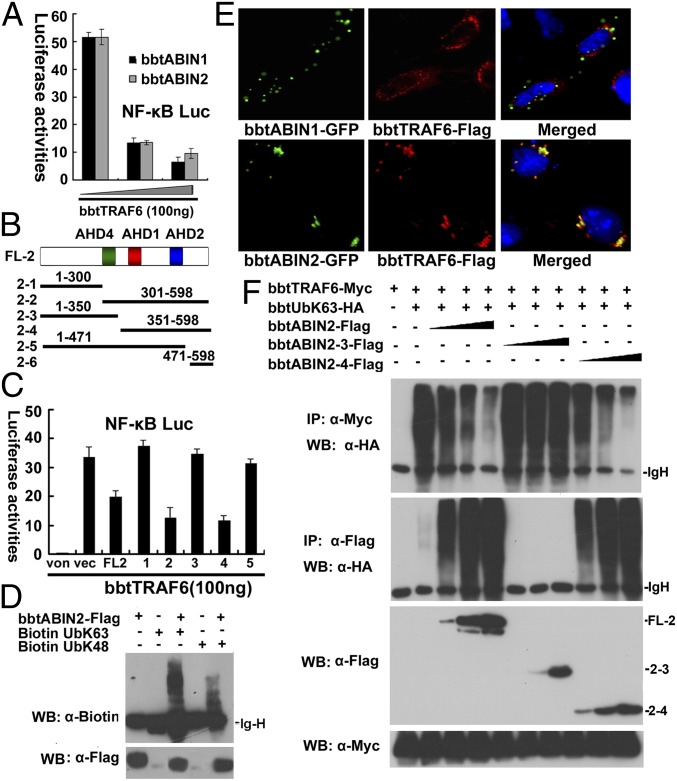
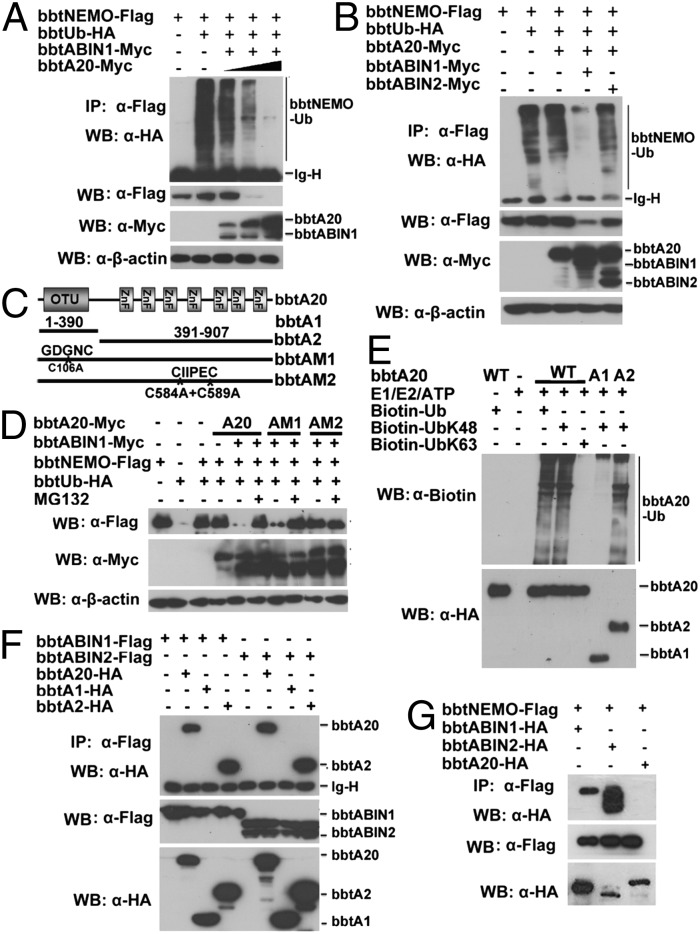
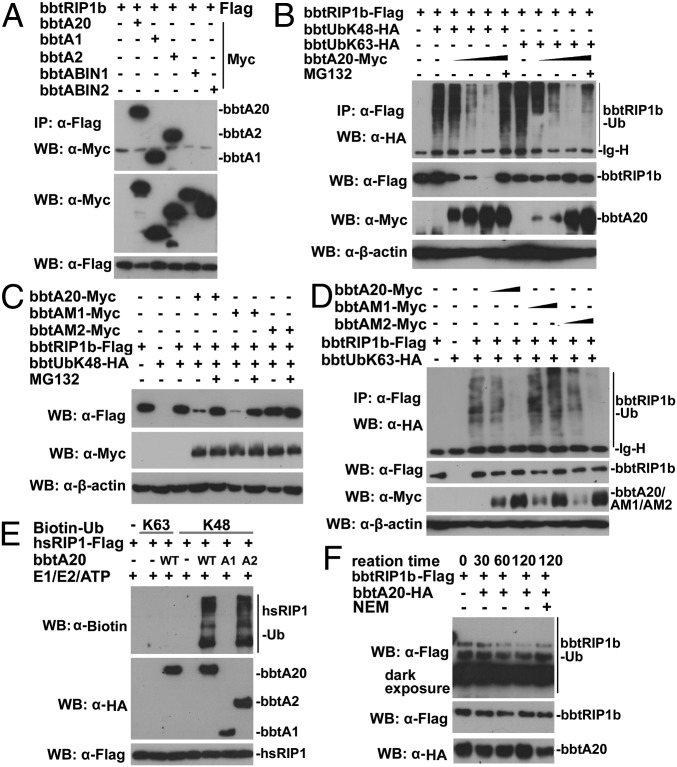
In the past decade, ubiquitination has been well documented to have multifaceted roles in regulating NF-κB activation in mammals. However, its function, especially how deubiquitinating enzymes balance the NF-κB activation, remains largely elusive in invertebrates. Investigating bbtA20 and its binding proteins, bbt A20-binding inhibitor of NF-κB (bbtABIN1) and bbtABIN2, in Chinese amphioxus Branchiostoma belcheri tsingtauense, we found that bbtABIN2 can colocalize and compete with bbt TNF receptor-associated factor 6 to connect the K63-linked polyubiquitin chains, whereas bbtABIN1 physically links bbtA20 to bbt NF-κB essential modulator (bbtNEMO) to facilitate the K48-linked ubiquitination of bbtNEMO. Similar to human A20, bbtA20 is a dual enzyme that removes the K63-linked polyubiquitin chains and builds the K48-linked polyubiquitin chains on bbt receptor-interacting serine/threonine protein kinase 1b, leading to the inhibition of NF-κB signaling. Our study not only suggests that ubiquitination is an ancient strategy in regulating NF-κB activation but also provides the first evidence, to our knowledge, for ABINs/A20-mediated inhibition of NF-κB via modifying the ubiquitinated proteins in a basal chordate, adding information on the stepwise development of vertebrate innate immune signaling.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
ABINs: A20 binding inhibitors of NF-kappa B and apoptosis signaling.Biochem Pharmacol. 2009 Jul 15;78(2):105-14. doi: 10.1016/j.bcp.2009.02.009. Epub 2009 Feb 27. Biochem Pharmacol. 2009. PMID: 19464428 Review.
-
The biology of A20-binding inhibitors of NF-kappaB activation (ABINs).Adv Exp Med Biol. 2014;809:13-31. doi: 10.1007/978-1-4939-0398-6_2. Adv Exp Med Biol. 2014. PMID: 25302363 Review.
-
Molecular basis for the unique deubiquitinating activity of the NF-kappaB inhibitor A20.J Mol Biol. 2008 Feb 15;376(2):526-40. doi: 10.1016/j.jmb.2007.11.092. Epub 2007 Dec 4. J Mol Biol. 2008. PMID: 18164316 Free PMC article.
-
Regulation of NF-κB signaling by the A20 deubiquitinase.Cell Mol Immunol. 2012 Mar;9(2):123-30. doi: 10.1038/cmi.2011.59. Epub 2012 Feb 20. Cell Mol Immunol. 2012. PMID: 22343828 Free PMC article. Review.
-
Identification of a novel A20-binding inhibitor of nuclear factor-kappa B activation termed ABIN-2.J Biol Chem. 2001 Aug 10;276(32):30216-23. doi: 10.1074/jbc.M100048200. Epub 2001 Jun 4. J Biol Chem. 2001. PMID: 11390377
Cited by
-
Current research into A20 mediation of allergic respiratory diseases and its potential usefulness as a therapeutic target.Front Immunol. 2023 Mar 28;14:1166928. doi: 10.3389/fimmu.2023.1166928. eCollection 2023. Front Immunol. 2023. PMID: 37056760 Free PMC article. Review.
-
Interleukin-17 signaling triggers degradation of the constitutive NF-κB inhibitor ABIN-1.Immunohorizons. 2017 Sep;1(7):133-141. doi: 10.4049/immunohorizons.1700035. Immunohorizons. 2017. PMID: 30761389 Free PMC article.
-
A20 Inhibits LPS-Induced Inflammation by Regulating TRAF6 Polyubiquitination in Rainbow Trout.Int J Mol Sci. 2021 Sep 10;22(18):9801. doi: 10.3390/ijms22189801. Int J Mol Sci. 2021. PMID: 34575978 Free PMC article.
-
Toll-Like Receptors, Associated Biological Roles, and Signaling Networks in Non-Mammals.Front Immunol. 2018 Jul 2;9:1523. doi: 10.3389/fimmu.2018.01523. eCollection 2018. Front Immunol. 2018. PMID: 30034391 Free PMC article. Review.
-
Expression and role of ABIN1 in sepsis: In vitro and in vivo studies.Open Med (Wars). 2020 Dec 4;16(1):33-40. doi: 10.1515/med-2021-0008. eCollection 2021. Open Med (Wars). 2020. Retraction in: Open Med (Wars). 2022 Jun 11;17(1):1064. doi: 10.1515/med-2022-0503. PMID: 33364432 Free PMC article. Retracted.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
