

Review
doi: 10.4161/cam.28437.
CCM1 and the second life of proteins in adhesion complexes
Affiliations
- PMID: 24714220
- PMCID: PMC4049860
- DOI: 10.4161/cam.28437
Item in Clipboard
Review
CCM1 and the second life of proteins in adhesion complexes
Cell Adh Migr.
2014.
Abstract
It is well recognized that a number of proteins present within adhesion complexes perform discrete signaling functions outside these adhesion complexes, including transcriptional control. In this respect, β-catenin is a well-known example of an adhesion protein present both in cadherin complexes and in the nucleus where it regulates the TCF transcription factor. Here we discuss nuclear functions of adhesion complex proteins with a special focus on the CCM-1/KRIT-1 protein, which may turn out to be yet another adhesion complex protein with a second life.
Keywords: CCM1/Krit-1; Rac Rho; adhesion signaling; nuclear translocation; small GTPases; transcription.
Figures
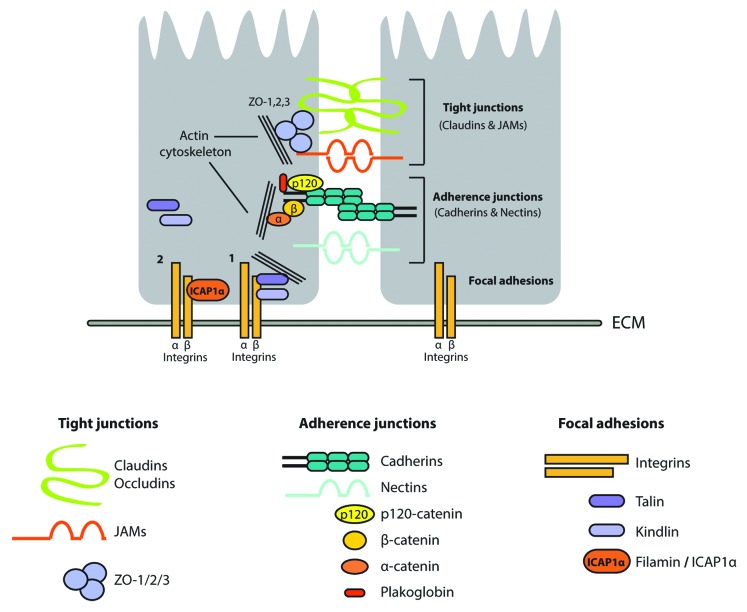
Figure 1. A simplified representation of the important mediators of cell–cell and cell–matrix adhesion. The adherens junctions consist of the Claudin and JAM families of transmembrane proteins, which are connected to the actin cytoskeleton via the ZO-family of proteins. Tight junctions consist of the catenin and nectin families of transmembrane proteins, which are connected to the cytoskeleton via the β-catenin interaction to α-catenin. Integrin-mediated cell–cell or cell–matrix interactions at focal adhesion sites is established via interaction of talins and kindlins to actin bundles (1). This can be inhibited by binding proteins such as filamin and ICAP1 to the β-integrin tail (2). For further details, see text.
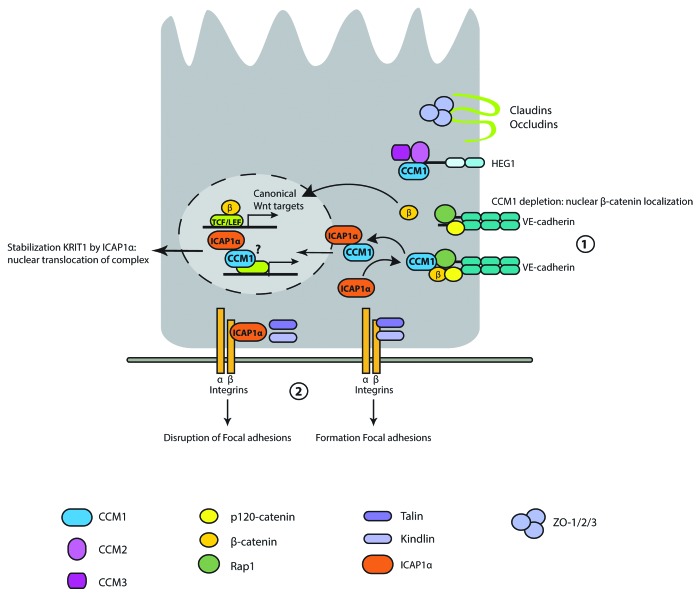
Figure 2. Molecular details of CCM1 biological function. Loss of CCM1 results in release of β-catenin from VE-cadherin, and subsequent activation of TCF/LEF-dependent transcription (1). Interaction of ICAP1 to β1-integrins disturbs focal adhesions by preventing binding of talin and kindlin. CCM1 inhibits binding of ICAP1 to β1-integrins and ICAP1 stabilizes CCM1 followed by nuclear translocation of the complex (2). CCM1 is located to the plasma membrane through interaction with the HEG1 transmembrane receptor.
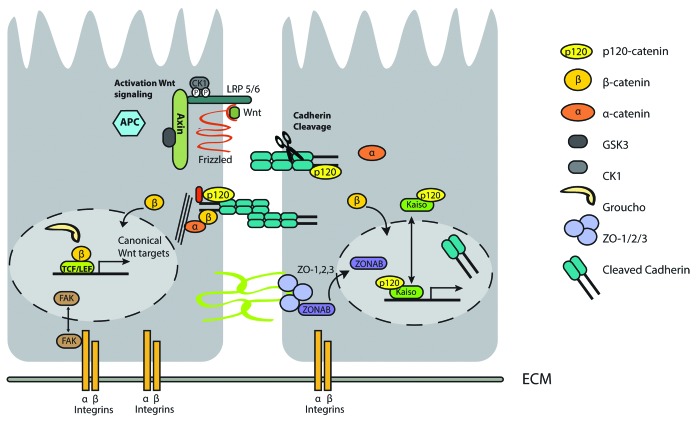
Figure 3. Dual role of proteins in adhesion complexes and transcription regulation. In absence of wnt signaling, β-catenin is degraded by the APC-destruction complex, whereas in presence of wnt signaling, degradation of β-catenin by APC is prevented and β-catenin activates TCF/LEF-mediated transcription (Left panel). Disruption of E-cadherin signaling by, for example, ADAM10-mediated cleavage of E-cadherin, results in nuclear translocation of β-catenin, and subsequent activation of the Wnt signaling pathway. However, following complete disintegration of the E-cadherin complex, the cytoplasmic domain of E-cadherin, derived after proteolytic cleavage and in addition p120ctn, may also translocate to the nucleus (Right panel). Combined or in isolation this nuclear shuttling of E-cadherin complex proteins may have different biological outcome. Multiple signals, like stress signals can result in nuclear localization of FAK, a key component of integrin signaling (left panel).
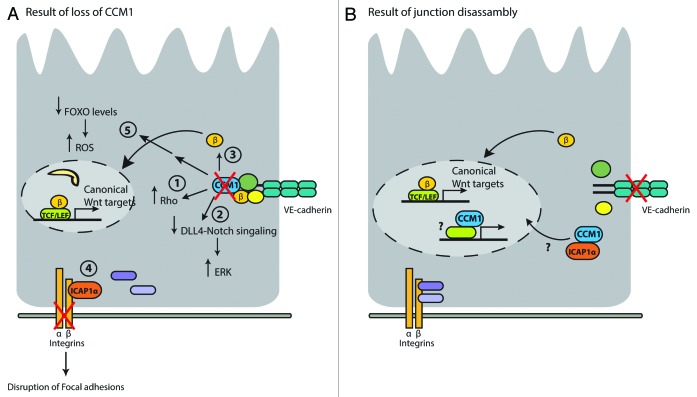
Figure 4. (A) Loss of CCM1 (depicted with a red cross in the figure) results in activation of Rho signaling (1), inhibition of DLL4-Notch signaling (2), nuclear localization of β-catenin (3), disruption of focal adhesions (indicated by red cross through integrins) as a result of ICAP binding (4), and decreased FOXO protein levels, resulting in increased ROS levels (5). (B) Suggested model in a situation where cadherin signaling is disrupted (depicted by a red cross through cadherin). See text for more details.
Similar articles
-
Rap1 and its effector KRIT1/CCM1 regulate beta-catenin signaling.Dis Model Mech. 2010 Jan-Feb;3(1-2):73-83. doi: 10.1242/dmm.003293. Epub 2009 Dec 9. Dis Model Mech. 2010. PMID: 20007487 Free PMC article.
-
CCM1 regulates vascular-lumen organization by inducing endothelial polarity.J Cell Sci. 2010 Apr 1;123(Pt 7):1073-80. doi: 10.1242/jcs.059329. J Cell Sci. 2010. PMID: 20332120
-
Wnt-5a-CKI{alpha} signaling promotes {beta}-catenin/E-cadherin complex formation and intercellular adhesion in human breast epithelial cells.J Biol Chem. 2009 Apr 17;284(16):10968-79. doi: 10.1074/jbc.M804923200. Epub 2009 Feb 25. J Biol Chem. 2009. PMID: 19244247 Free PMC article.
-
Recent insights into cerebral cavernous malformations: a complex jigsaw puzzle under construction.FEBS J. 2010 Mar;277(5):1084-96. doi: 10.1111/j.1742-4658.2009.07537.x. Epub 2010 Jan 22. FEBS J. 2010. PMID: 20096036 Free PMC article. Review.
-
Coordinating Rho and Rac: the regulation of Rho GTPase signaling and cadherin junctions.Prog Mol Biol Transl Sci. 2013;116:49-68. doi: 10.1016/B978-0-12-394311-8.00003-0. Prog Mol Biol Transl Sci. 2013. PMID: 23481190 Review.
Cited by
-
Molecular analysis of gastric cancer identifies genomic markers of drug sensitivity in Asian gastric cancer.J Cancer. 2018 Jul 30;9(16):2973-2980. doi: 10.7150/jca.25506. eCollection 2018. J Cancer. 2018. PMID: 30123366 Free PMC article.
-
Molecular Genetic Features of Cerebral Cavernous Malformations (CCM) Patients: An Overall View from Genes to Endothelial Cells.Cells. 2021 Mar 22;10(3):704. doi: 10.3390/cells10030704. Cells. 2021. PMID: 33810005 Free PMC article. Review.
-
Cancer-secreted exosomal miR-21-5p induces angiogenesis and vascular permeability by targeting KRIT1.Cell Death Dis. 2021 Jun 4;12(6):576. doi: 10.1038/s41419-021-03803-8. Cell Death Dis. 2021. PMID: 34088891 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous