

Review
doi: 10.1101/cshperspect.a018366.
Regulation of the X chromosomes in Caenorhabditis elegans
Affiliations
- PMID: 24591522
- PMCID: PMC3942922
- DOI: 10.1101/cshperspect.a018366
Item in Clipboard
Review
Regulation of the X chromosomes in Caenorhabditis elegans
Cold Spring Harb Perspect Biol.
.
Abstract
Dosage compensation, which regulates the expression of genes residing on the sex chromosomes, has provided valuable insights into chromatin-based mechanisms of gene regulation. The nematode Caenorhabditis elegans has adopted various strategies to down-regulate and even nearly silence the X chromosomes. This article discusses the different chromatin-based strategies used in somatic tissues and in the germline to modulate gene expression from the C. elegans X chromosomes and compares these strategies to those used by other organisms to cope with similar X-chromosome dosage differences.
Figures
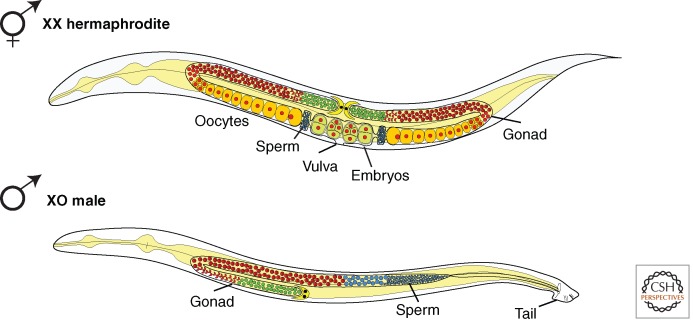
C. elegans hermaphrodite and male anatomy. C. elegans naturally exists as two sexes: XX hermaphrodites and XO males. Hermaphrodites and males display several sex-specific anatomical features, most notably a male tail designed for mating and a vulva on the ventral surface of hermaphrodites for reception of male sperm and for egg-laying. Their germline programs also differ. The two-armed gonad in hermaphrodites produces sperm initially and then oocytes throughout adulthood. The one-armed gonad in males produces sperm continuously. (Adapted, with permission, from Hansen et al. 2004, © Elsevier.)
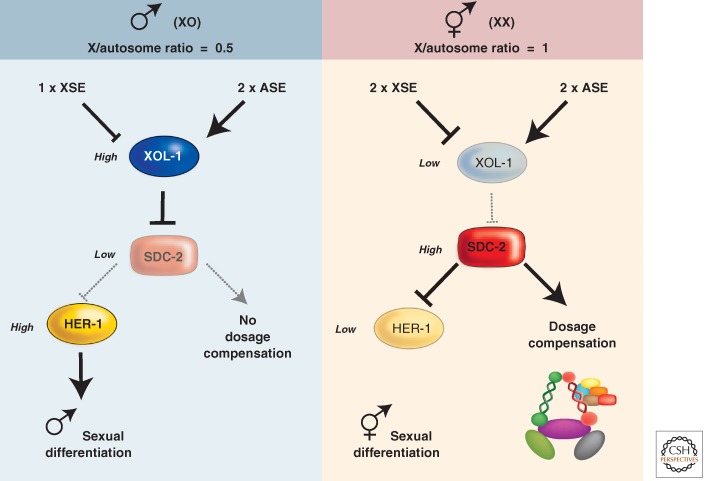
Pathway of sex determination and dosage compensation in C. elegans. The proposed roles of X signal elements (XSEs) and autosomal signal elements (ASEs) in regulating XOL-1 levels and the subsequent sexual differentiation and assembly of the dosage compensation complex (DCC), which represses X-linked gene expression about twofold in XX hermaphrodites, are highlighted.
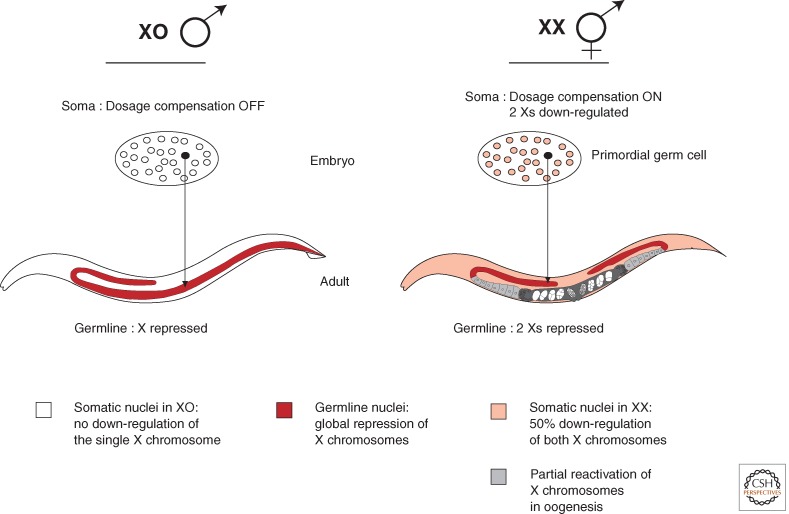
Overview of X-chromosome regulation. Dosage compensation occurs in somatic tissues only in XX hermaphrodites. Repression of the Xs in the germline occurs in both XO males and XX hermaphrodites. Hermaphrodites display late and partial activation of X-linked genes during late pachytene of oogenesis. The black cell highlights the single primordial germ cell in the embryo that generates the germline in the adult gonad.
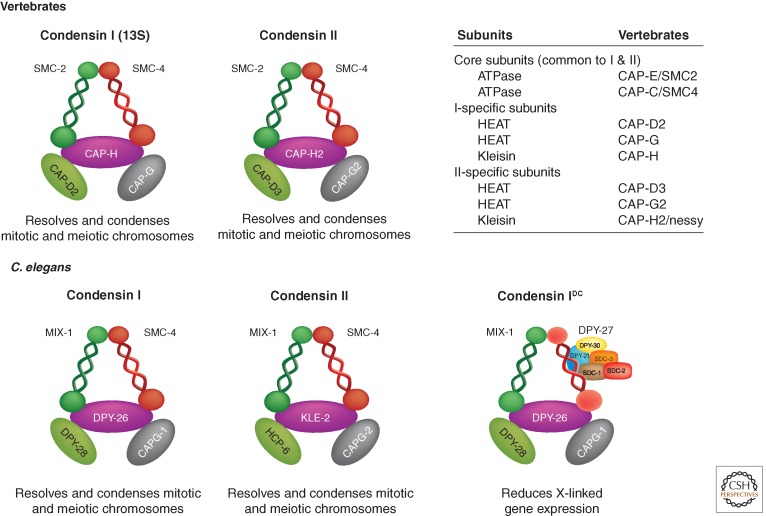
The DCC and condensin complexes. The worm DCC resembles the condensin complex, which functions in condensing chromosomes during nuclear division. In particular, the DCC contains several subunits that are homologous to the XCAP (XCAP for Xenopus chromosome-associated polypeptide) subunits of the 13S condensin complex I, originally characterized in Xenopus. There are two condensin complexes in most metazoans and three complexes in C. elegans. MIX-1 is present in all three C. elegans condensin complexes. Three additional DCC subunits (DPY-26, DPY-28, and CAPG-1) are present in both condensin I and condensin IDC. The SDC proteins, DPY-21 and DPY-30, do not resemble known condensin subunits; they instead function in localizing condensin IDC to the X chromosome. (Adapted from Meyer 2005 and Csankovszki et al. 2009.)
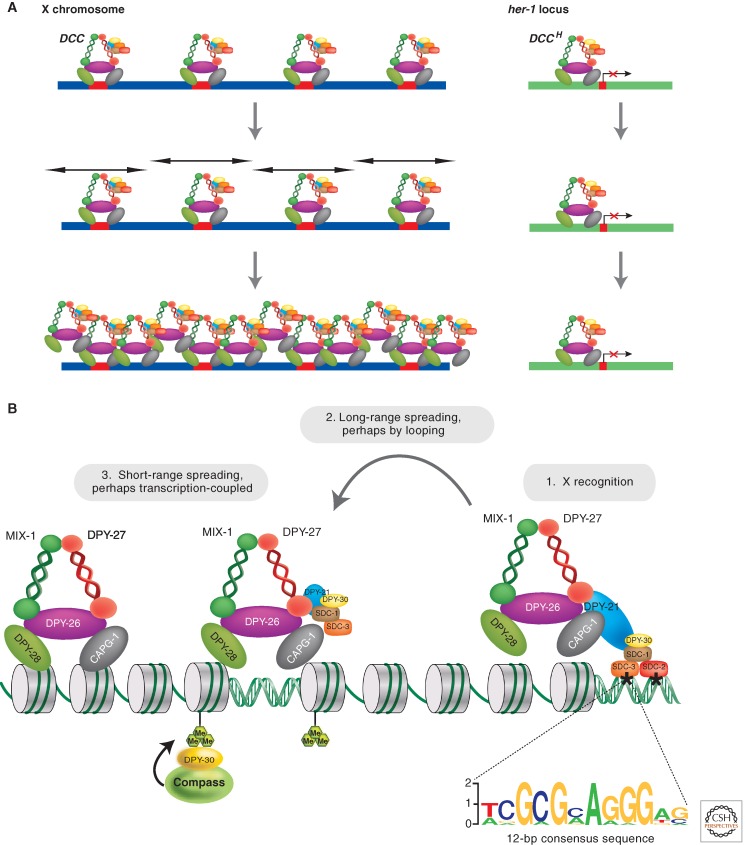
DCC recruitment and spreading on the X chromosome. (A) The DCC is recruited to approximately 100 to 300 recruitment elements and spreads along both X chromosomes. It also binds to the upstream region of the autosomal gene her-1 and reduces expression by 20-fold, denoted DCCH to indicate that the complex that binds to her-1 lacks the DPY-21 protein. (Adapted from Alekseyenko and Kuroda 2004.) (B) From initial recruitment sites, the DCC spreads and accumulates preferentially at promoters by an unknown mechanism. The recruitment sites on the X chromosome, indicated by asterisks, contain multiple 12-bp DNA sequence motifs (shown below the DNA) that are important for recruitment. DCC binding is dynamic during development and is tuned to the transcriptional activity of individual genes. DCC accumulation is high in the body of highly expressed genes. (Adapted from Ercan and Lieb 2009.)
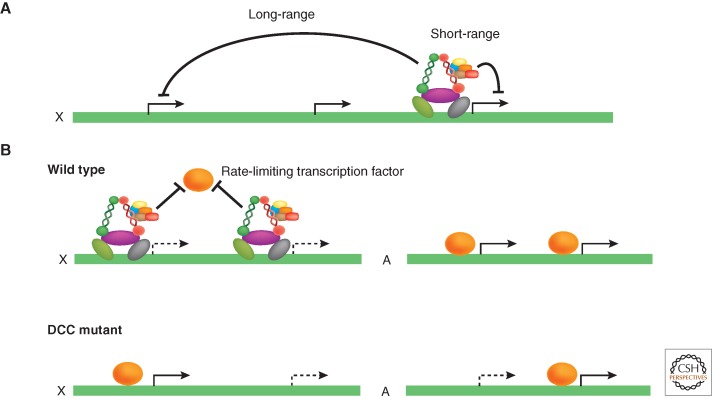
Models for how DCC concentrated on the X chromosomes may regulate gene expression. Upon loss of DCC function, most of the gene expression changes on the X chromosome are an increase in transcription, consistent with the DCC repressing X-chromosome transcription. However, only about half of DCC-bound genes increase in expression and many genes whose expression increases are not bound by the DCC. (A) The DCC may repress genes locally or cause structural changes that affect distant loci. (B) On the autosomes, the majority of expression changes due to loss of DCC function are a decrease in transcription. This could be explained by a model in which the DCC repels an activator from the X chromosomes. In the absence of the DCC, the activator is more evenly distributed between the X chromosomes and autosomes, resulting in increased gene expression from the X and decreased gene expression from the autosomes. An alternative model (not shown) is that the DCC may repress genes locally, and in the absence of the DCC additional effects on the X and autosomes result from the increase in transcription of hundreds of genes encoded from the X. The possibilities discussed above are not mutually exclusive.
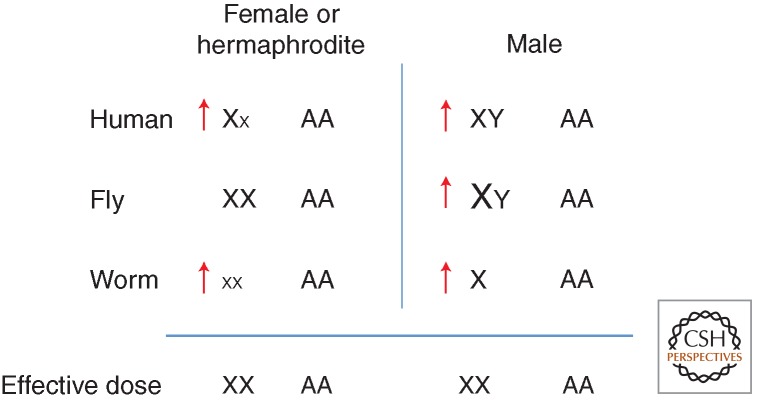
Coordinated up-regulation and DCC-mediated down-regulation of X-linked gene expression. Dosage compensation between the sexes is accomplished differently in the three well-studied systems shown. In mammals, one of the X chromosomes in females is inactivated (small x). In fruit flies, transcription from the single X in males is increased by a factor of 2 (larger X). In worms, transcription from both of the X chromosomes in hermaphrodites is decreased by a factor of 2 (smaller Xs). In all three systems, transcript levels from the X chromosome(s) and autosomes are similar in both females/hermaphrodites and males, suggesting that there is a mechanism to increase X transcription approximately twofold in both sexes (red arrows).
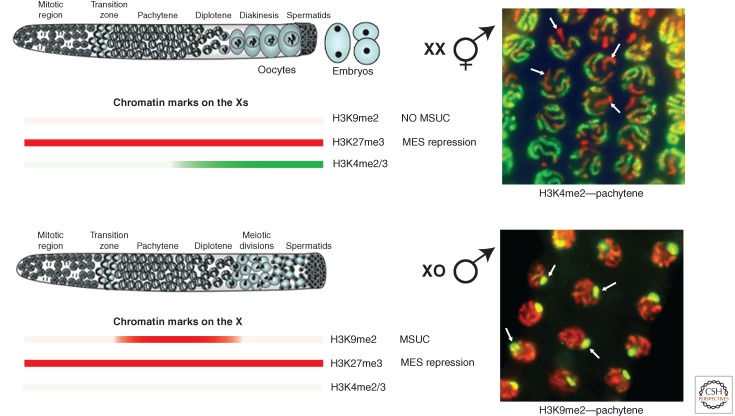
Epigenetic regulation of the X chromosomes during germ cell development. In both sexes, germ cells progress through mitosis (left), enter meiosis in the transition zone, and progress through meiosis I prophase. Cells destined to form sperm in both sexes complete the meiotic divisions in the gonad. In hermaphrodites, cells destined to form oocytes progress through meiotic prophase in the gonad and complete the meiotic divisions after ovulation and fertilization. The presence of various histone modifications on the X chromosome(s) in germ cells is shown by red bars (for repressive modifications) and green bars (for activating modifications). As shown on the right, antibodies to particular histone modifications reveal that the X chromosomes in germ nuclei are marked differently than the autosomes and are repressed. H3K4me2 (green), a mark of actively expressed chromatin, is excluded from the Xs in XX pachytene nuclei. H3K9me2 (green), a mark of heterochromatin, is concentrated on the X in XO pachytene nuclei as part of MSUC (meiotic silencing of unsynapsed chromatin). DNA is stained red. Arrows indicate representative X chromosomes in each image.
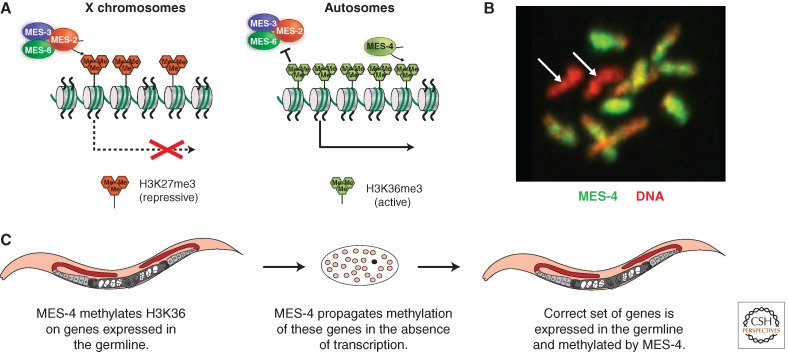
Model for the transgenerational role of MES-4 and how MES-4 and MES-2/3/6 participate in X repression in the germline. (A) MES-2/3/6-generated H3K27me3, a repressive histone modification, is concentrated on the X chromosomes. MES-4 and H3K36 methylation on autosomal genes repel the MES-2/3/6 complex, helping to concentrate its repressive action on the X chromosomes. (B) MES-4 is concentrated on the autosomes. MES-4 immunostaining is in green. DNA is stained red. Arrows mark the two X chromosomes, which lack MES-4 staining. (C) MES-4 associates with genes expressed in the maternal germline and marks them with H3K36me2/3. MES-4 propagation of H3K36 methyl marks in the absence of transcription enables MES-4 to transmit the memory of germline gene expression across generations.
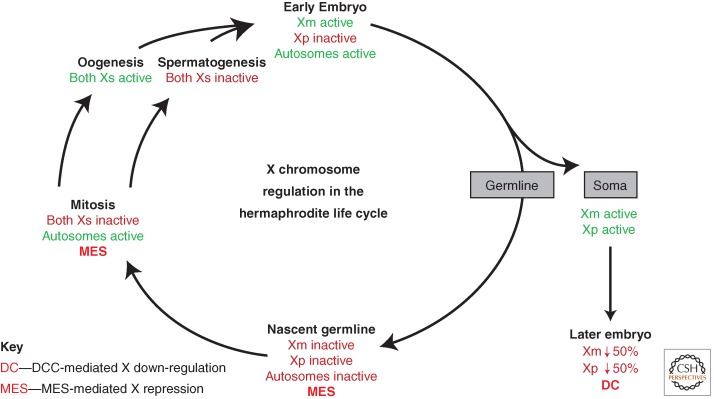
Regulation of the X chromosomes through the life cycle of an XX hermaphrodite. The X chromosomes are regulated by different mechanisms at different stages and in different tissues: paternal X inactivation in the early embryo, dosage compensation (DC) in the somatic tissues of 30-cell and later-stage embryos and worms, and MES-mediated repression in the germline. In XO male meiosis (not depicted), meiotic silencing of the unsynapsed X by MSUC additionally occurs in the germline.
Similar articles
-
Restricting dosage compensation complex binding to the X chromosomes by H2A.Z/HTZ-1.PLoS Genet. 2009 Oct;5(10):e1000699. doi: 10.1371/journal.pgen.1000699. Epub 2009 Oct 23. PLoS Genet. 2009. PMID: 19851459 Free PMC article.
-
Untangling the Contributions of Sex-Specific Gene Regulation and X-Chromosome Dosage to Sex-Biased Gene Expression in Caenorhabditis elegans.Genetics. 2016 Sep;204(1):355-69. doi: 10.1534/genetics.116.190298. Epub 2016 Jun 29. Genetics. 2016. PMID: 27356611 Free PMC article.
-
A molecular link between gene-specific and chromosome-wide transcriptional repression.Genes Dev. 2002 Apr 1;16(7):796-805. doi: 10.1101/gad.972702. Genes Dev. 2002. PMID: 11937488 Free PMC article.
-
X-Chromosome dosage compensation.WormBook. 2005 Jun 25:1-14. doi: 10.1895/wormbook.1.8.1. WormBook. 2005. PMID: 18050416 Free PMC article. Review.
-
C. elegans dosage compensation: a window into mechanisms of domain-scale gene regulation.Chromosome Res. 2009;17(2):215-27. doi: 10.1007/s10577-008-9011-0. Chromosome Res. 2009. PMID: 19308702 Review.
Cited by
-
Dynamic Control of X Chromosome Conformation and Repression by a Histone H4K20 Demethylase.Cell. 2017 Sep 21;171(1):85-102.e23. doi: 10.1016/j.cell.2017.07.041. Epub 2017 Aug 31. Cell. 2017. PMID: 28867287 Free PMC article.
-
Status of dosage compensation of X chromosome in bovine genome.Genetica. 2016 Aug;144(4):435-44. doi: 10.1007/s10709-016-9912-3. Epub 2016 Jul 4. Genetica. 2016. PMID: 27376899
-
Regulatory Divergence as a Mechanism for X-Autosome Incompatibilities in Caenorhabditis Nematodes.Genome Biol Evol. 2023 Apr 6;15(4):evad055. doi: 10.1093/gbe/evad055. Genome Biol Evol. 2023. PMID: 37014784 Free PMC article.
-
Polycomb repressive complex's evolutionary conserved function: the role of EZH2 status and cellular background.Clin Epigenetics. 2016 May 27;8:55. doi: 10.1186/s13148-016-0226-1. eCollection 2016. Clin Epigenetics. 2016. PMID: 27239242 Free PMC article. Review.
-
Repression of somatic cell fate in the germline.Cell Mol Life Sci. 2015 Oct;72(19):3599-620. doi: 10.1007/s00018-015-1942-y. Epub 2015 Jun 5. Cell Mol Life Sci. 2015. PMID: 26043973 Free PMC article. Review.
References
-
- Alekseyenko AA, Kuroda MI 2004. Molecular biology. Filling gaps in genome organization. Science 303: 1148–1149 - PubMed
WWW RESOURCE
-
- http://www.wormbook.org/chapters/www_dosagecomp/dosagecomp.html Meyer BJ 2005. X-chromosome dosage compensation. In WormBook. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources