

Glial wingless/Wnt regulates glutamate receptor clustering and synaptic physiology at the Drosophila neuromuscular junction
- PMID: 24553932
- PMCID: PMC3931504
- DOI: 10.1523/JNEUROSCI.3714-13.2014
Glial wingless/Wnt regulates glutamate receptor clustering and synaptic physiology at the Drosophila neuromuscular junction
Abstract
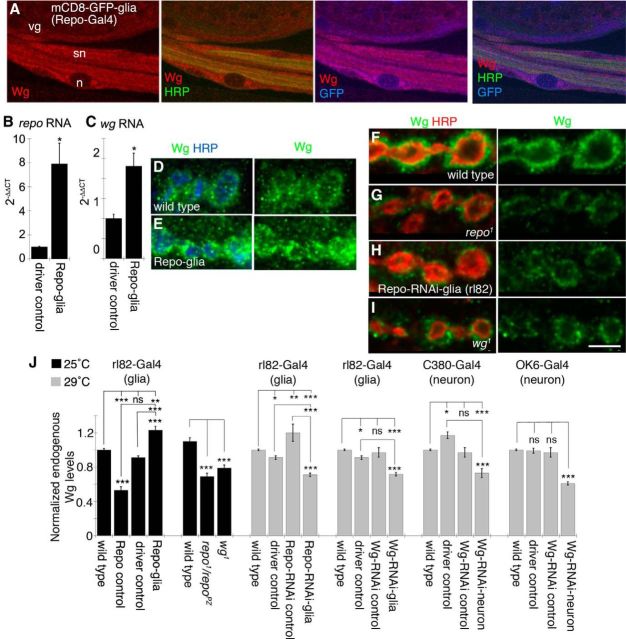
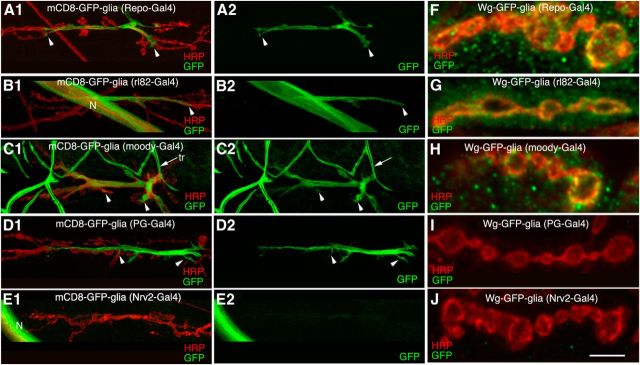
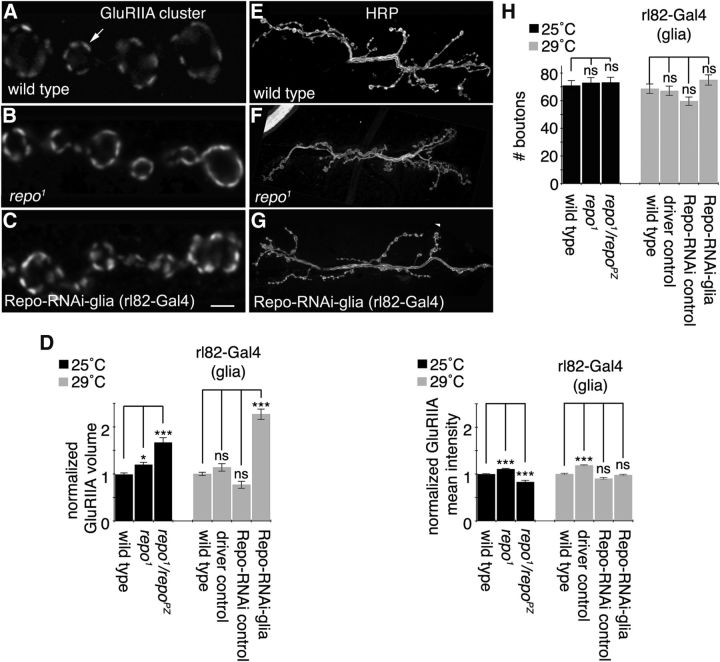
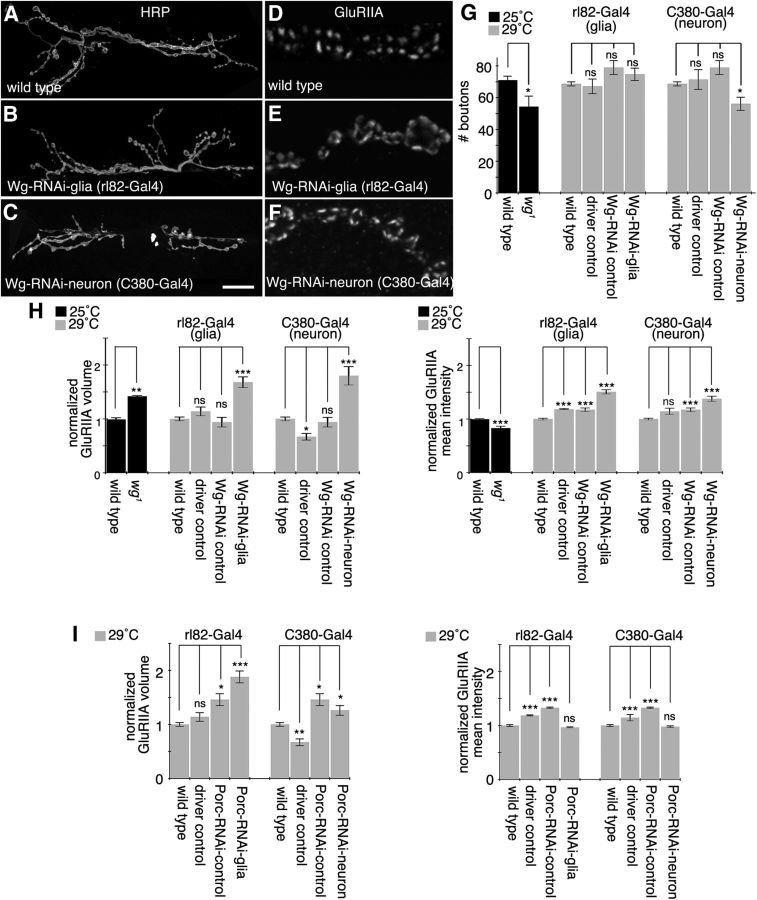
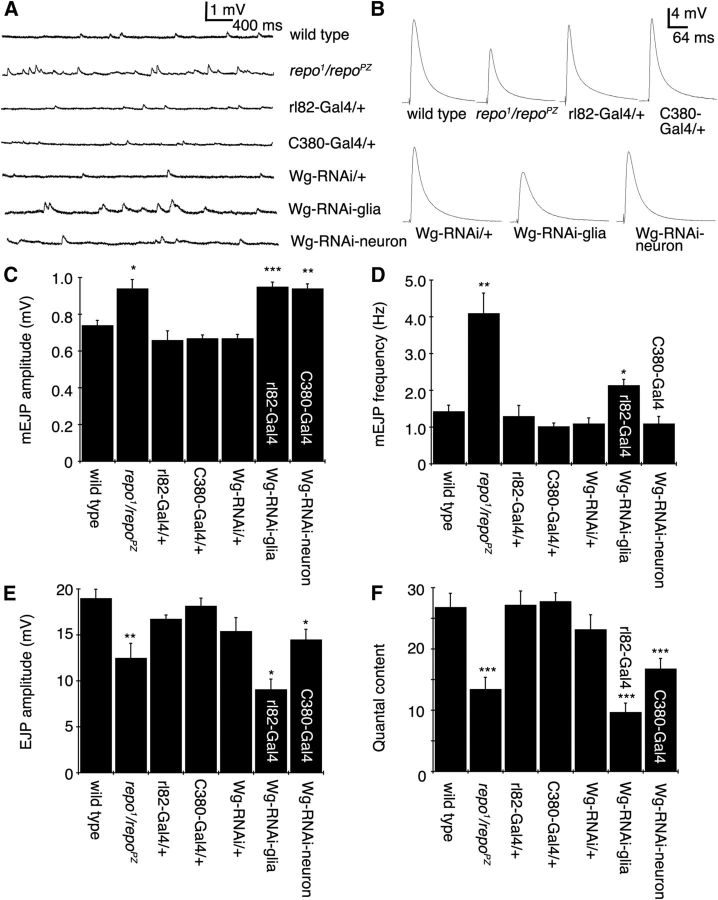
Glial cells are emerging as important regulators of synapse formation, maturation, and plasticity through the release of secreted signaling molecules. Here we use chromatin immunoprecipitation along with Drosophila genomic tiling arrays to define potential targets of the glial transcription factor Reversed polarity (Repo). Unexpectedly, we identified wingless (wg), a secreted morphogen that regulates synaptic growth at the Drosophila larval neuromuscular junction (NMJ), as a potential Repo target gene. We demonstrate that Repo regulates wg expression in vivo and that local glial cells secrete Wg at the NMJ to regulate glutamate receptor clustering and synaptic function. This work identifies Wg as a novel in vivo glial-secreted factor that specifically modulates assembly of the postsynaptic signaling machinery at the Drosophila NMJ.
Keywords: Drosophila; NMJ; glia; synapse; wnt/Wg.
Figures

Similar articles
-
Postsynaptic glutamate receptors regulate local BMP signaling at the Drosophila neuromuscular junction.Development. 2014 Jan;141(2):436-47. doi: 10.1242/dev.097758. Epub 2013 Dec 18. Development. 2014. PMID: 24353060 Free PMC article.
-
Kismet positively regulates glutamate receptor localization and synaptic transmission at the Drosophila neuromuscular junction.PLoS One. 2014 Nov 20;9(11):e113494. doi: 10.1371/journal.pone.0113494. eCollection 2014. PLoS One. 2014. PMID: 25412171 Free PMC article.
-
Carrier of Wingless (Cow) Regulation of Drosophila Neuromuscular Junction Development.eNeuro. 2020 Mar 10;7(2):ENEURO.0285-19.2020. doi: 10.1523/ENEURO.0285-19.2020. Print 2020 Mar/Apr. eNeuro. 2020. PMID: 32024666 Free PMC article.
-
Development and plasticity of the Drosophila larval neuromuscular junction.Wiley Interdiscip Rev Dev Biol. 2013 Sep-Oct;2(5):647-70. doi: 10.1002/wdev.108. Epub 2013 Feb 5. Wiley Interdiscip Rev Dev Biol. 2013. PMID: 24014452 Free PMC article. Review.
-
Receptor clustering: nothing succeeds like success.Curr Biol. 2004 Jun 8;14(11):R413-5. doi: 10.1016/j.cub.2004.05.031. Curr Biol. 2004. PMID: 15182686 Review.
Cited by
-
Revisiting the role of the Gcm transcription factor, from master regulator to Swiss army knife.Fly (Austin). 2016 Oct;10(4):210-8. doi: 10.1080/19336934.2016.1212793. Epub 2016 Jul 19. Fly (Austin). 2016. PMID: 27434165 Free PMC article.
-
Epigenetic Signaling in Glia Controls Presynaptic Homeostatic Plasticity.Neuron. 2020 Feb 5;105(3):491-505.e3. doi: 10.1016/j.neuron.2019.10.041. Epub 2019 Dec 3. Neuron. 2020. PMID: 31810838 Free PMC article.
-
Signal Exchange through Extracellular Vesicles in Neuromuscular Junction Establishment and Maintenance: From Physiology to Pathology.Int J Mol Sci. 2019 Jun 8;20(11):2804. doi: 10.3390/ijms20112804. Int J Mol Sci. 2019. PMID: 31181747 Free PMC article. Review.
-
Brain-specific lipoprotein receptors interact with astrocyte derived apolipoprotein and mediate neuron-glia lipid shuttling.Nat Commun. 2021 Apr 23;12(1):2408. doi: 10.1038/s41467-021-22751-7. Nat Commun. 2021. PMID: 33893307 Free PMC article.
-
Primary Cilia in Glial Cells: An Oasis in the Journey to Overcoming Neurodegenerative Diseases.Front Neurosci. 2021 Sep 30;15:736888. doi: 10.3389/fnins.2021.736888. eCollection 2021. Front Neurosci. 2021. PMID: 34658775 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
