

The association of mitochondrial potential and copy number with pig oocyte maturation and developmental potential
- PMID: 24492657
- PMCID: PMC3999391
- DOI: 10.1262/jrd.2013-098
The association of mitochondrial potential and copy number with pig oocyte maturation and developmental potential
Abstract
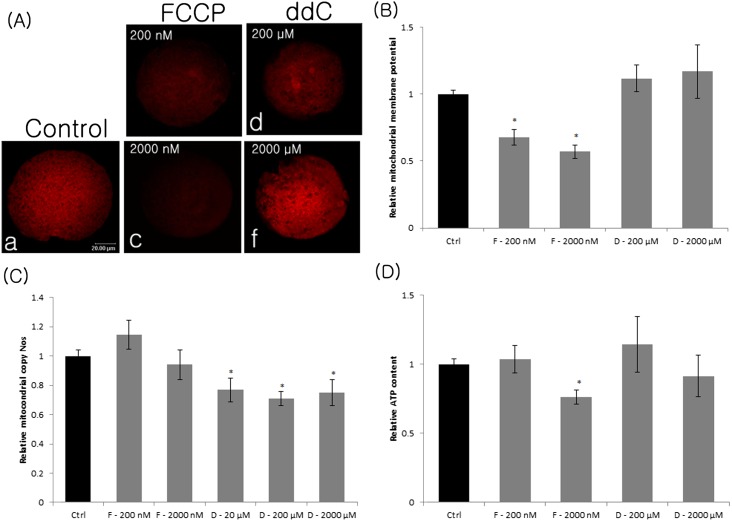
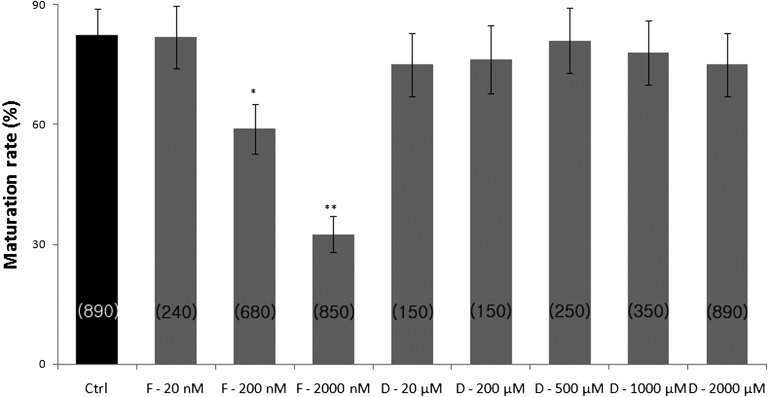
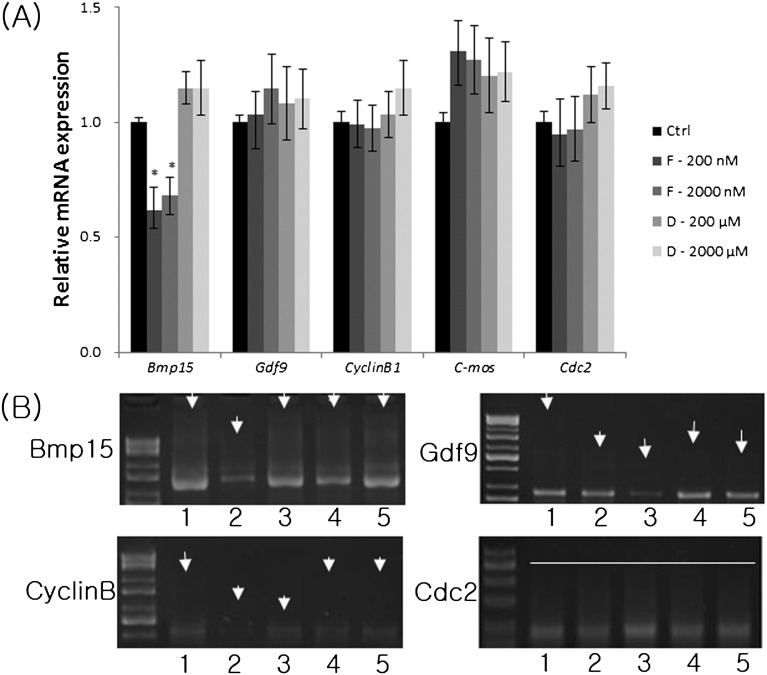
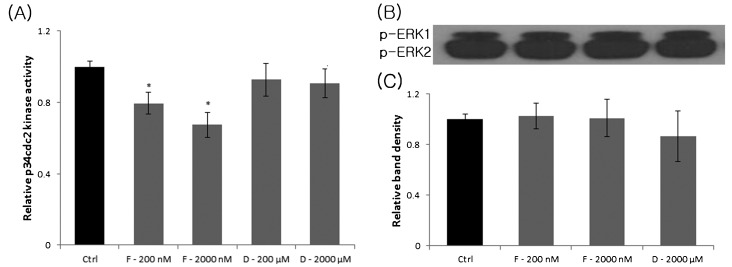
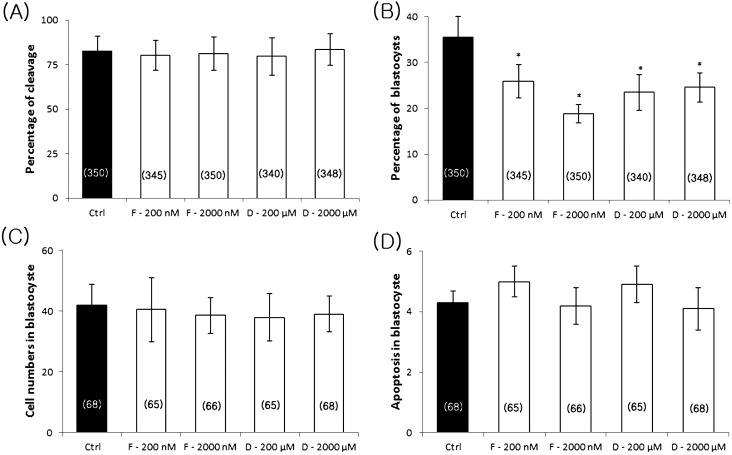
ATP is critical for oocyte maturation, fertilization, and subsequent embryo development. Both mitochondrial membrane potential and copy number expand during oocyte maturation. In order to differentiate the roles of mitochondrial metabolic activity and mtDNA copy number during oocyte maturation, we used two inhibitors, FCCP (carbonyl cyanide p-(tri-fluromethoxy)phenyl-hydrazone) and ddC (2'3-dideoxycytidine), to deplete the mitochondrial membrane potential (Δφm) and mitochondrial copy number, respectively. FCCP (2000 nM) reduced ATP production by affecting mitochondrial Δφm, decreased the mRNA expression of Bmp15 (bone morphogenetic protein 15), and shortened the poly(A) tails of Bmp15, Gdf9 (growth differentiation factor 9), and Cyclin B1 transcripts. FCCP (200 and 2000 nM) also affected p34(cdc2) kinase activity. By contrast, ddC did not alter ATP production. Instead, ddC significantly decreased mtDNA copy number (P < 0.05). FCCP (200 and 2000 nM) also decreased extrusion of the first polar body, whereas ddC at all concentrations did not affect the ability of immature oocytes to reach metaphase II. Both FCCP (200 and 2000 nM) and ddC (200 and 2000 µM) reduced parthenogenetic blastocyst formation compared with untreated oocytes. However, these inhibitors did not affect total cell number and apoptosis. These findings suggest that mitochondrial metabolic activity is critical for oocyte maturation and that both mitochondrial metabolic activity and replication contribute to the developmental competence of porcine oocytes.
Figures
Similar articles
-
The importance of mitochondrial metabolic activity and mitochondrial DNA replication during oocyte maturation in vitro on oocyte quality and subsequent embryo developmental competence.Mol Reprod Dev. 2012 Jun;79(6):392-401. doi: 10.1002/mrd.22042. Epub 2012 Apr 16. Mol Reprod Dev. 2012. PMID: 22467220
-
Regulated mitochondrial DNA replication during oocyte maturation is essential for successful porcine embryonic development.Biol Reprod. 2007 Feb;76(2):327-35. doi: 10.1095/biolreprod.106.054536. Epub 2006 Oct 11. Biol Reprod. 2007. PMID: 17035641
-
Regulation of maternal gene expression by MEK/MAPK and MPF signaling in porcine oocytes during in vitro meiotic maturation.J Reprod Dev. 2011 Feb;57(1):49-56. doi: 10.1262/jrd.10-087h. Epub 2010 Sep 10. J Reprod Dev. 2011. PMID: 20834195
-
Mitochondrial function in the human oocyte and embryo and their role in developmental competence.Mitochondrion. 2011 Sep;11(5):797-813. doi: 10.1016/j.mito.2010.09.012. Epub 2010 Oct 7. Mitochondrion. 2011. PMID: 20933103 Review.
-
[Mitochondrial and oocyte development].Yi Chuan. 2007 Dec;29(12):1429-33. Yi Chuan. 2007. PMID: 18065375 Review. Chinese.
Cited by
-
Spatiotemporal Distribution and Function of Mitochondria in Oocytes.Reprod Sci. 2024 Feb;31(2):332-340. doi: 10.1007/s43032-023-01331-8. Epub 2023 Aug 21. Reprod Sci. 2024. PMID: 37605038 Review.
-
Fe(III) Is Essential for Porcine Embryonic Development via Mitochondrial Function Maintenance.PLoS One. 2015 Jul 10;10(7):e0130791. doi: 10.1371/journal.pone.0130791. eCollection 2015. PLoS One. 2015. PMID: 26161974 Free PMC article.
-
Effect of Endometriosis on Cumulus ATP, Number of Mitochondria and Oocyte Maturity in Cumulus Oocyte Complex in Mice.Rev Bras Ginecol Obstet. 2023 Jul;45(7):e393-e400. doi: 10.1055/s-0043-1772186. Epub 2023 Aug 18. Rev Bras Ginecol Obstet. 2023. PMID: 37595596 Free PMC article.
-
Evaluation of mitochondria in mouse oocytes following cisplatin exposure.J Ovarian Res. 2021 May 10;14(1):65. doi: 10.1186/s13048-021-00817-w. J Ovarian Res. 2021. PMID: 33971923 Free PMC article.
-
The Molecular Quality and Mitochondrial Activity of Porcine Cumulus-Oocyte Complexes Are Affected by Their Exposure to Three Endocrine-Active Compounds under 3D In Vitro Maturation Conditions.Int J Mol Sci. 2022 Apr 20;23(9):4572. doi: 10.3390/ijms23094572. Int J Mol Sci. 2022. PMID: 35562963 Free PMC article.
References
-
- Stojkovic M, Machado SA, Stojkovic P, Zakhartchenko V, Hutzler P, Gonçalves PB, Wolf E. Mitochondrial distribution and adenosine triphosphate content of bovine oocytes before and after in vitro maturation: correlation with morphological criteria and developmental capacity after in vitro fertilization and culture. Biol Reprod 2001; 64: 904–909 - PubMed
-
- Nagano M, Katagiri S, Takahashi Y. Relationship between bovine oocyte morphology and in vitro developmental potential. Zygote 2006; 14: 53–61 - PubMed
-
- Santos TA, El Shourbagy S, St John JC. Mitochondrial content reflects oocyte variability and fertilization outcome. Fertil Steril 2006; 85: 584–591 - PubMed
-
- Reynier P, May-Panloup P, Chrétien MF, Morgan CJ, Jean M, Savagner F, Barrière P, Malthièry Y. Mitochondrial DNA content affects the fertilizability of human oocytes. Mol Hum Reprod 2001; 7: 425–429 - PubMed
-
- Pikó L, Matsumoto L. Number of mitochondria and some properties of mitochondrial DNA in the mouse egg. Dev Biol 1976; 49: 1–10 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous