

Review
doi: 10.1021/cr4004709.
Epub 2014 Jan 29.
Radical S-adenosylmethionine enzymes
Affiliations
- PMID: 24476342
- PMCID: PMC4002137
- DOI: 10.1021/cr4004709
Item in Clipboard
Review
Radical S-adenosylmethionine enzymes
Chem Rev.
.
No abstract available
Figures
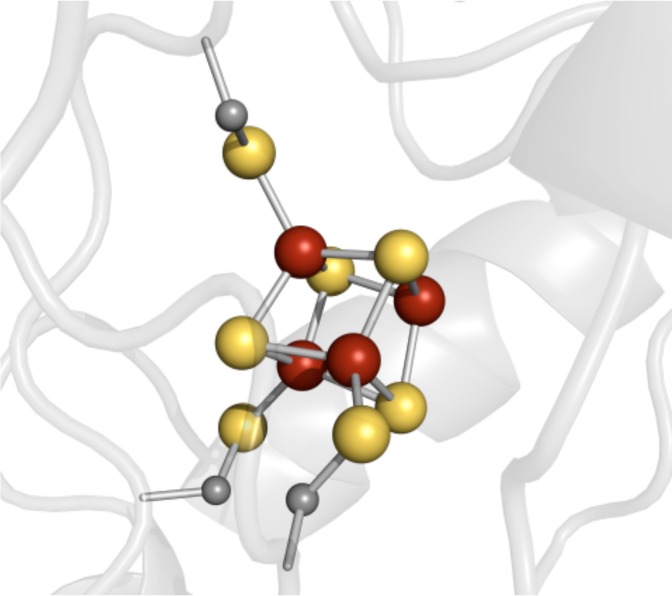
The site-differentiated [4Fe–4S] cluster coordinated
by
the CX3CX2C radical SAM motif (PDB ID 3IIZ).
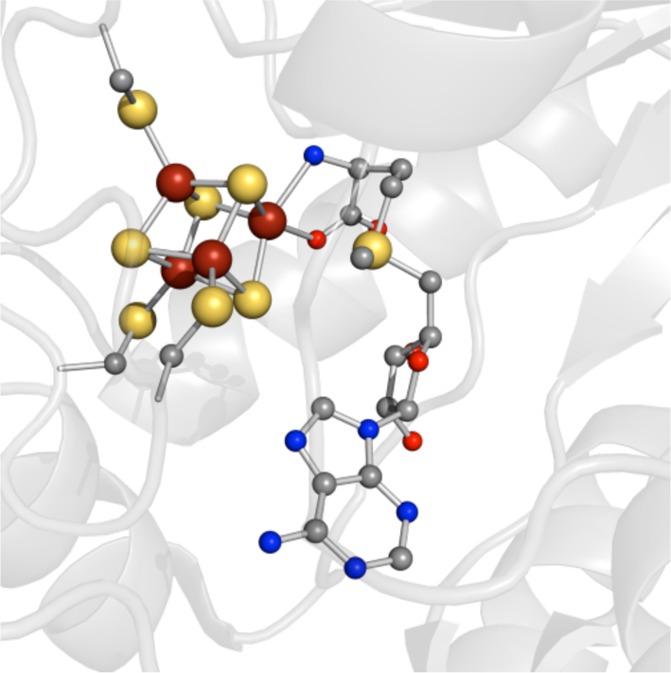
The bidentate coordination
of S-adenosylmethionine
to the unique iron site of the [4Fe–4S] cluster in radical
SAM enzymes (PDB ID 3IIZ).
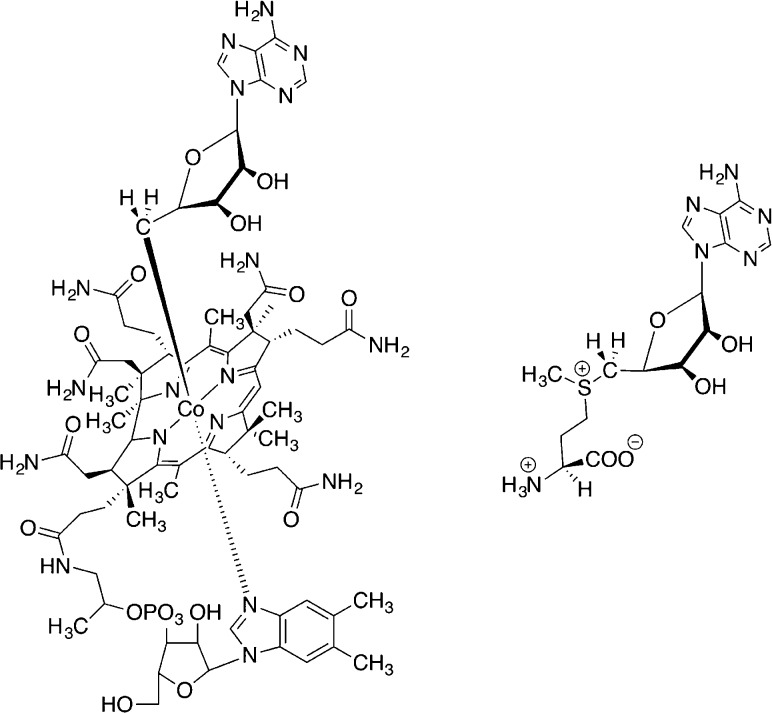
Structures of AdoCbl (left) and SAM (right).
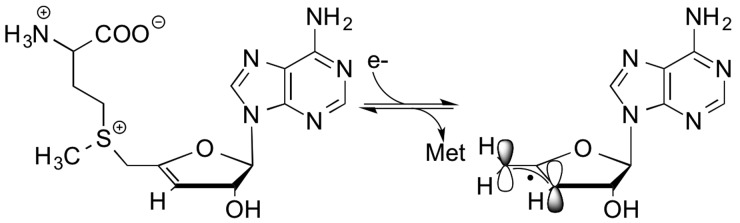
Reductive cleavage of S-3′,4′-anhydroadenosyl-l -methionine (anSAM) results in generation of the stable allylic
radical species 5′-deoxy-3′,4′-anhydroadenosine-5′-yl
(anAdo•).
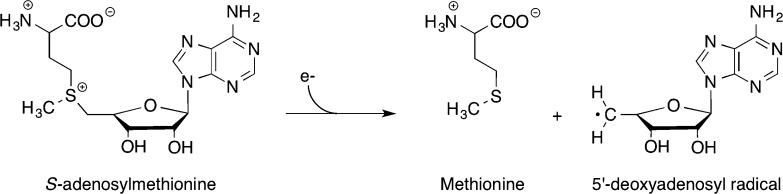
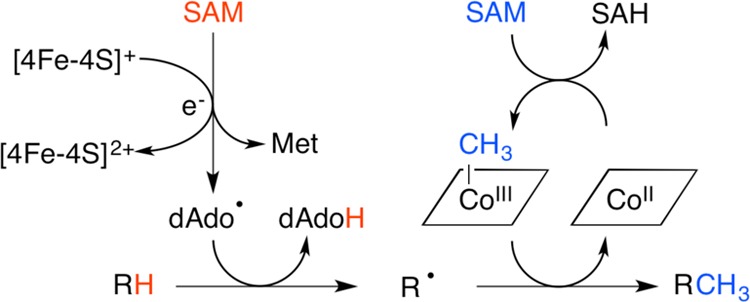
The cleavage of SAM to
generate methionine and the dAdo• is a reductive
cleavage event, requiring the input of one electron.
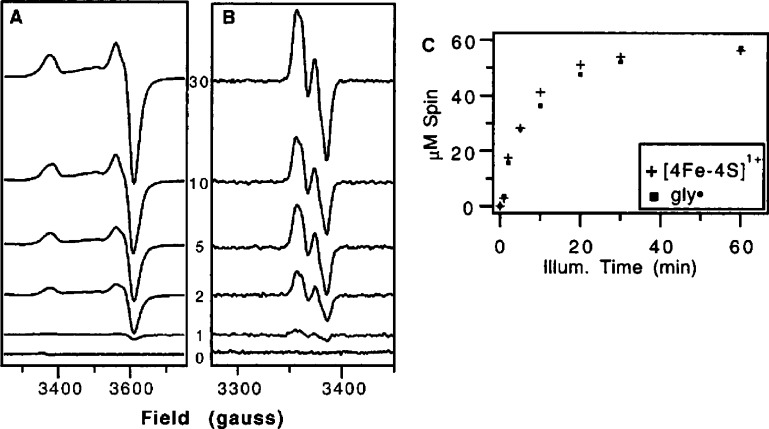
X-band EPR spectra of photoreduced PFL-AE before (A) and
after
addition of PFL (B), photoreduction time indicated in minutes. (C)
Spin quantitation of the EPR spectra in (A) for the amount of the
[4Fe–4S]+ cluster (+) and EPR spectra in (B) for
the amount of glycyl radical (■) as a function of illumination
time. Reprinted with permission from ref (21). Copyright 2000 American Chemical Society.
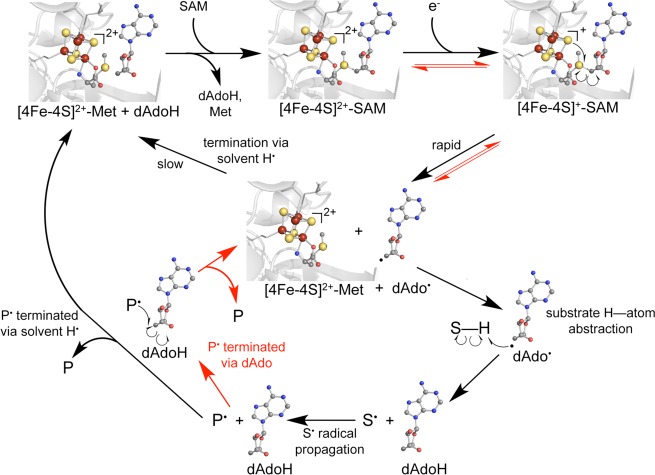
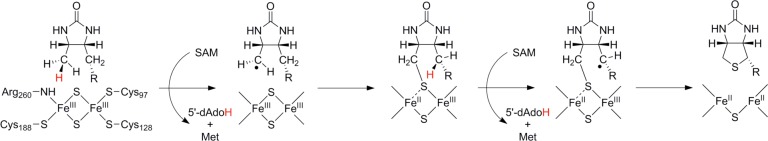
Framework mechanism for radical SAM cleavage
(PDB ID 3IIZ).
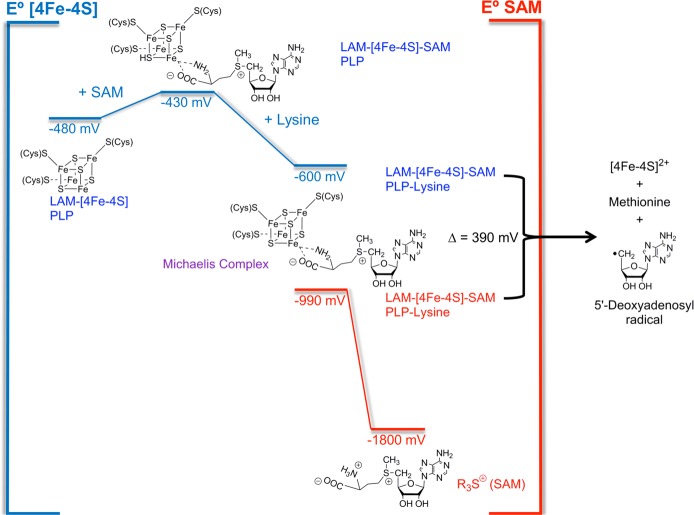
Reduction
potentials for SAM and the [4Fe–4S] cluster based
on experimental measurements for LAM.
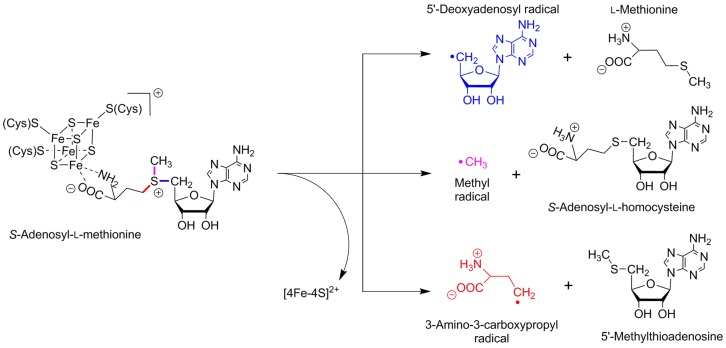
Regioselective
cleavage of the S–C bonds of SAM. Bonds that
may undergo enzymatic-based homolytic cleavage are demarked in varying
colors with S–C(5′) in blue, S–C(γ) in
red, and the S–C(methyl) in magenta.
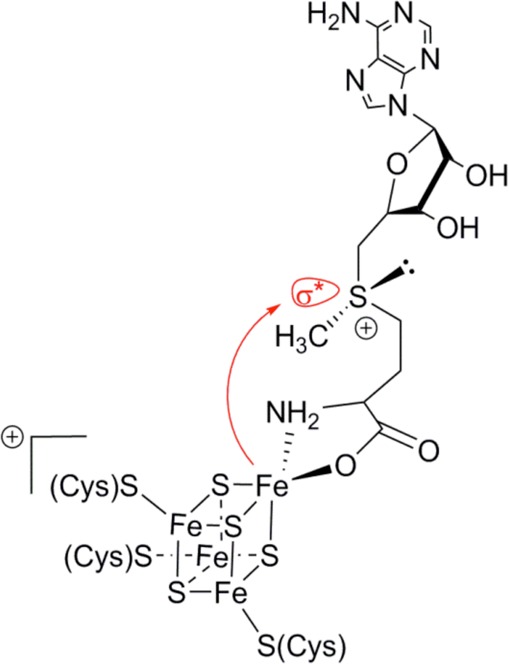
The reductive
cleavage of SAM occurs through an inner-sphere mechanism
involving a direct path between the unique iron ion of the cluster
and the sulfonium group antibonding S–C(5′) orbital.
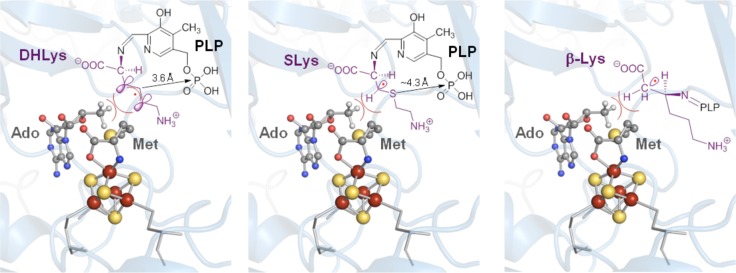
Illustration of the results of the LAM ENDOR
studies using stabilized
substrate and product radical analogue intermediates (PDB ID 2A5H). In all cases,
van der Waals contacts are maintained between the 5′-methyl
of dAdoH (carbons shown in gray) and the substrate/product radicals.
Illustrations for the substrate radicals generated upon reaction with trans-4,5-dehydro- l -lysine (DHLys, left), 4-thia-l -lysine (SLys, middle), and the product radical generated upon
equilibration of the reduced state of the enzyme with SAM and l -α-lysine (right). Adapted with permission from ref (38). Copyright 2006 American
Chemical Society.
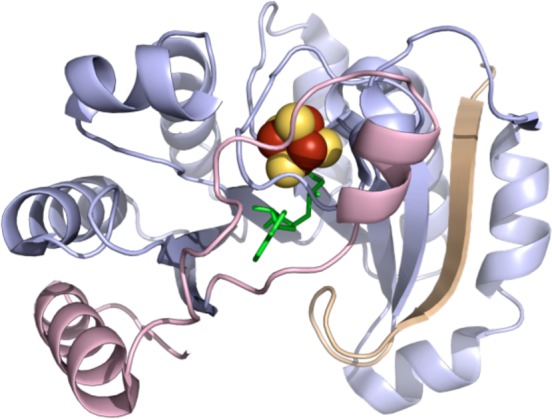
Example of a radical
SAM partial TIM barrel structure (PDB ID 3CB8 for PFL-AE). N-terminal
domain colored in wheat, radical SAM domain in light blue, C-terminal
domain in light pink, [4Fe–4S] cluster in yellow and rust spheres,
and SAM in green sticks.
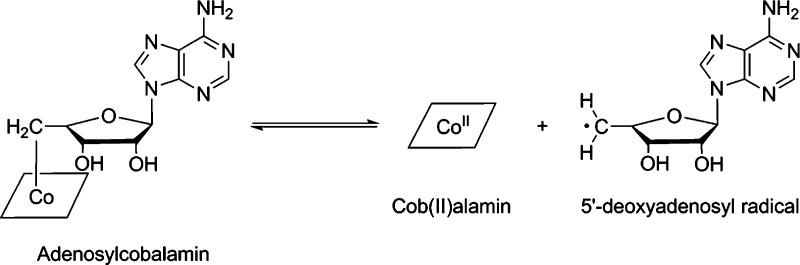
Homolytic cleavage of the Co–C bond to generate
cob(II)alamin
and the 5′- deoxyadenosyl radical.
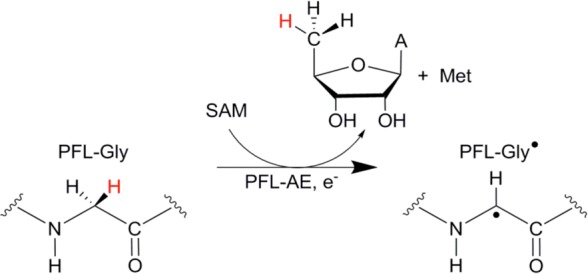
PFL-AE reaction
scheme catalyzing the activation of PFL by stereospecific
(pro-S) hydrogen atom abstraction
from PFL G734 (in E. coli).
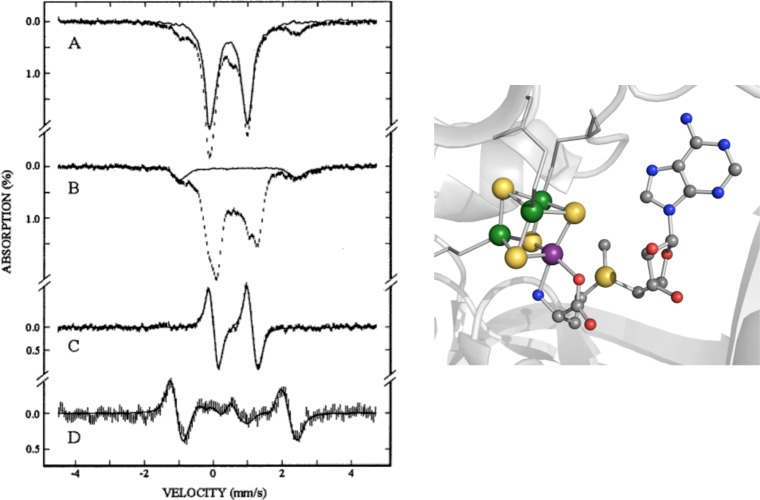
Left: Mössbauer spectra of 56Fe PFL-AE reconstituted
with 57Fe for incorporation into the unique iron site in
the absence (A) and presence (B) of SAM. The solid line in (A) is
the experimental spectrum of [4Fe–4S]2+ clusters
in PFL-AE normalized to 70% of the total Fe absorption of (A). The
solid line in (B) is the spectrum of the control sample containing
only the reconstitution ingredients and SAM but without PFL-AE and
is normalized to 15% of the total Fe absorption of (B). A difference
spectrum of (B) minus (A) is shown in (C). Spectrum (D) is a difference
spectrum of the samples (A) and (B) recorded in a parallel field of
8 T. Reprinted with permission from ref (93a). Copyright 2002 American chemical Society.
Right: Illustration of the PFL-AE [4Fe–4S] cluster with 57Fe (purple sphere) in the unique site bound by SAM, with
the other sites occupied by natural abundance iron (56Fe,
green spheres) (PDB ID 3CB8).
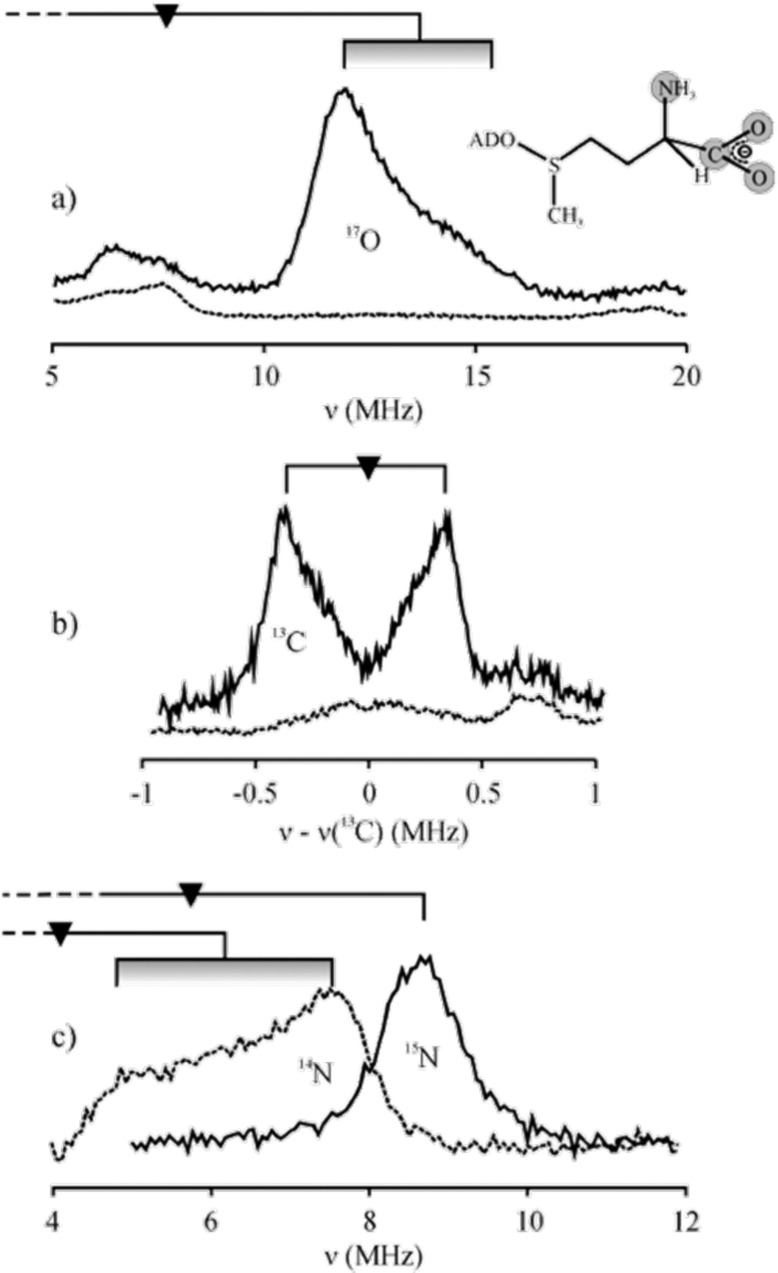
35-GHz pulsed ENDOR spectra of PFL-AE with 17O (A) and 13C (B) carboxylato-labeled and 15N-amino-labeled
(C) SAM as compared to data from an unlabeled sample, at g⊥. Reprinted with permission from ref (67). Copyright 2005 American
Chemical Society.
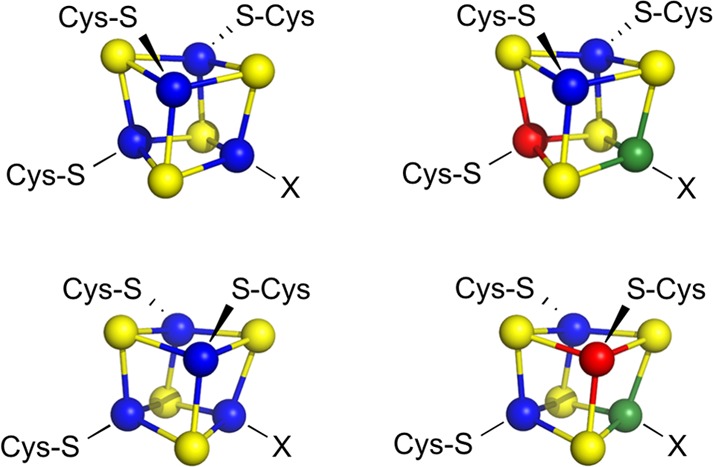
Representative
valence delocalization of biological [4Fe–4S]2+ clusters
containing two Fe2.5+–Fe2.5+ pairs (left,
top and bottom). Representative valence localization
of [4Fe–4S]2+ clusters in PFL-AE isolated from whole
cells containing one Fe2.5+–Fe2.5+ pair
and one Fe3+–Fe2+ pair (right, top and
bottom) (PDB ID 3CB8).
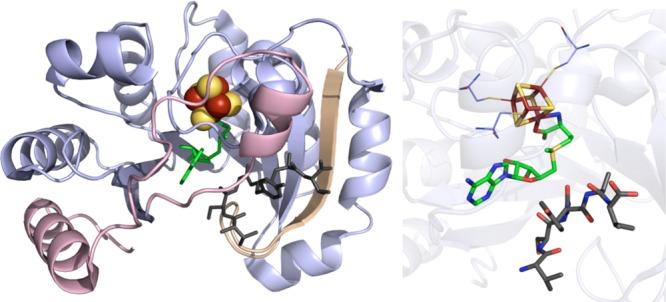
PFL-AE crystal structure
(PDB ID 3CB8). Left: N-terminal domain colored in
wheat, radical SAM domain in light blue, C-terminal domain in light
pink, [4Fe–4S] cluster in yellow and rust spheres, SAM in green
sticks, 7-mer peptide in dark gray sticks. Right: Active site of PFL-AE
where [4Fe–4S] cluster (yellow and rust), SAM (green carbons),
and 7-mer peptide (gray carbons) are depicted in sticks with oxygens
colored red and nitrogens colored blue. Cysteines (light blue carbons)
involved in ligating cluster are depicted in lines.
Docking model of PFL-AE (PDB ID 3CB8). Best dock as produced
by ZDOCK, with
Cα of G734 in spacefill and radical domain of PFL (residues
712–759) in magenta. PFL-AE helices in cyan, strands in yellow,
and loops in gray. The [4Fe–4S] cluster (yellow and rust) and
SAM (green carbons) are depicted in sticks with oxygens colored red
and nitrogens colored blue. Reprinted with permission from ref (29). Copyright 2008 National
Academy of Sciences.
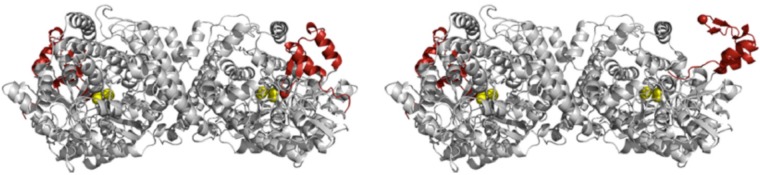
Schematic
representation of PFL: left, crystal structure of closed
conformation (PDB ID 2PFL); and right, model for the open conformation. Radical domain shown
in red where Gly734 is a red sphere and active site residues Cys418
and Cys419 are yellow spheres. Reprinted with permission from ref (149). Copyright 2010 American
Society for Biochemistry and Molecular Biology.
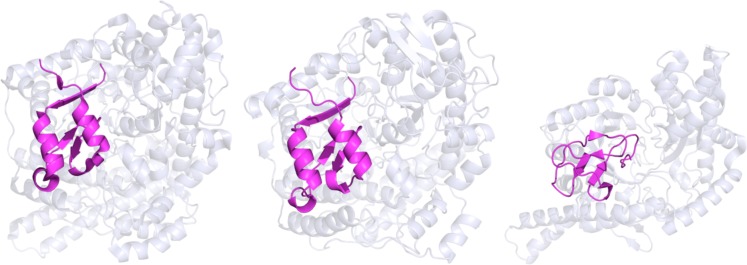
The X-ray crystal structures
of the activating enzyme substrates
from left to right: GD, PFL, and aRNR (PDB IDs: 1R8W, 2PFL, and 1HK8, respectively).
All structures possess a core 10-stranded β-barrel motif assembled
in a manner antiparallel to two parallel five-stranded β-sheets.
The β-barrel core is surrounded by α-helices forming the
β/α-barrel. Radical domains, highlighted in magenta, for
GD, PFL, and aRNR are composed of the amino acids 731–782,
702–754, and 540–586 (where aRNR possesses a mostly
disordered C-terminal domain), respectively.
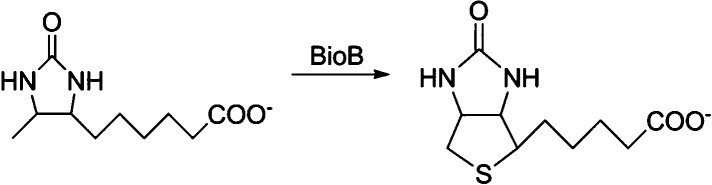
The conversion of dethiobiotin to biotin catalyzed by
biotin synthase
(BioB).
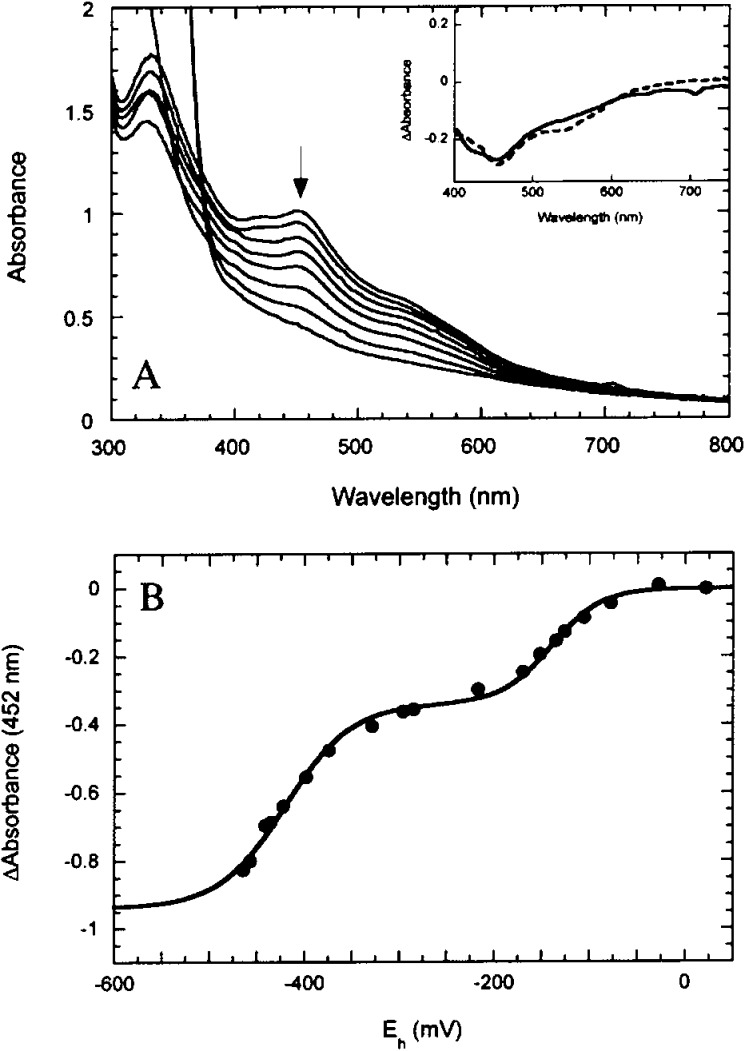
Reduction of BioB containing [2Fe–2S]2+ clusters.
(A) UV/visible spectra of BioB were recorded as the cell potential
was lowered by titration with dithionite. (Inset) Difference spectra
associated with the first wave of reduction (solid curve) and the
second wave of reduction (dashed curve) having maxima at 460 nm. (B)
The absorbance change at 452 nm was followed as a function of the
measured cell potential. Reprinted with permission from ref (24a). Copyright 2001 American
Chemical Society.
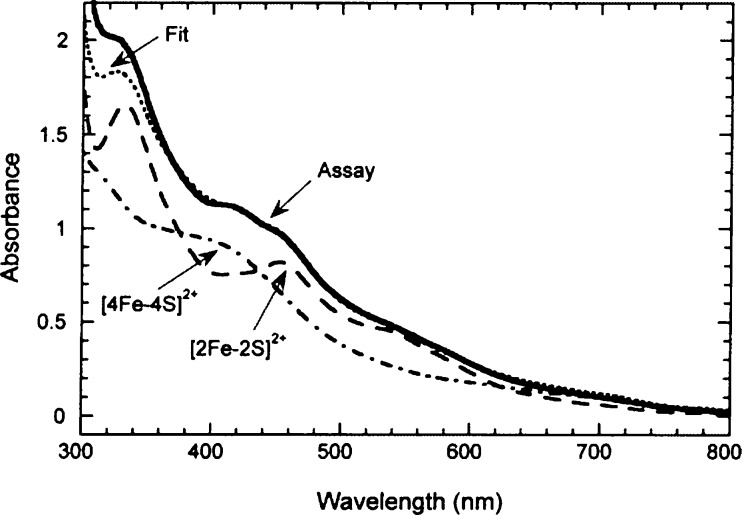
UV–visible
spectrum of BioB under assay conditions reveals
features characteristic of both [4Fe–4S]2+ and [2Fe–2S]2+ clusters. Reprinted with permission from ref (24a). Copyright 2001 American
Chemical Society.
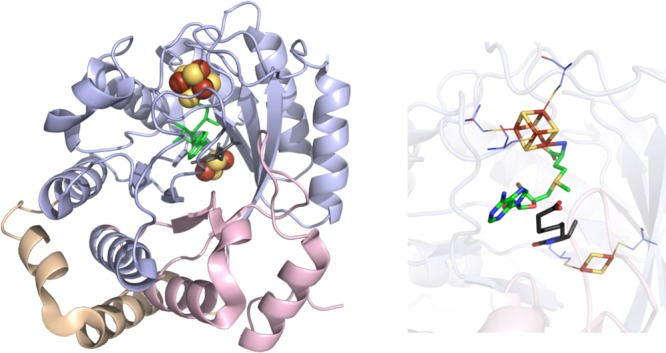
BioB crystal structure (PDB ID 1R30). Left: N-terminal domain colored in
wheat, radical SAM domain in light blue, C-terminal domain in light
pink, [4Fe–4S] and [2Fe–2S] clusters in yellow and rust
spheres, SAM in green sticks, dethiobiotin in dark gray sticks. Right:
Active site of BioB where [4Fe–4S] and [2Fe–2S] clusters
(yellow and rust), SAM (green carbons), and dethiobiotin (gray carbons)
are depicted in sticks with oxygens colored red and nitrogens colored
blue. Cysteines (light blue carbons) involved in ligating clusters
are depicted in lines.
The mechanism of biotin formation from dethiobiotin as catalyzed
by BioB.
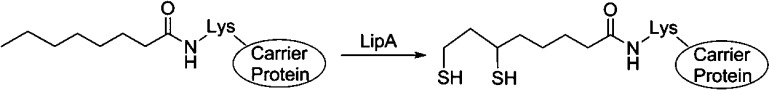
LipA reaction scheme
catalyzing the conversion of octanoyl-acyl
carrier protein to lipoyl-acyl carrier protein.
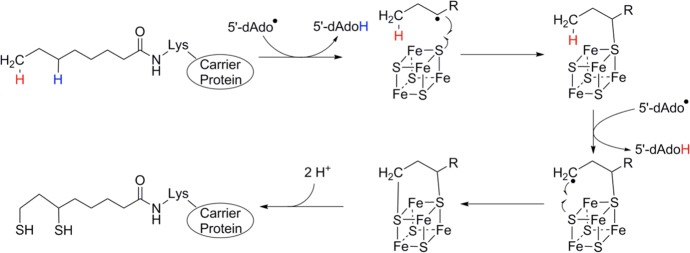
The mechanism of lipoyl-acyl carrier protein from octanoyl-acyl
carrier protein as catalyzed by LipA.
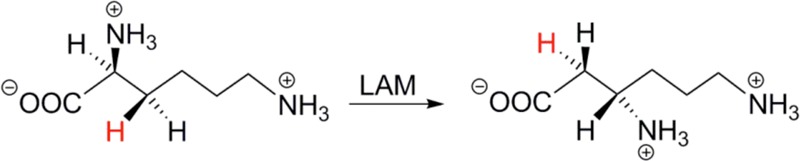
LAM reaction
scheme catalyzing the conversion of l -α-lysine
to l -β-lysine.
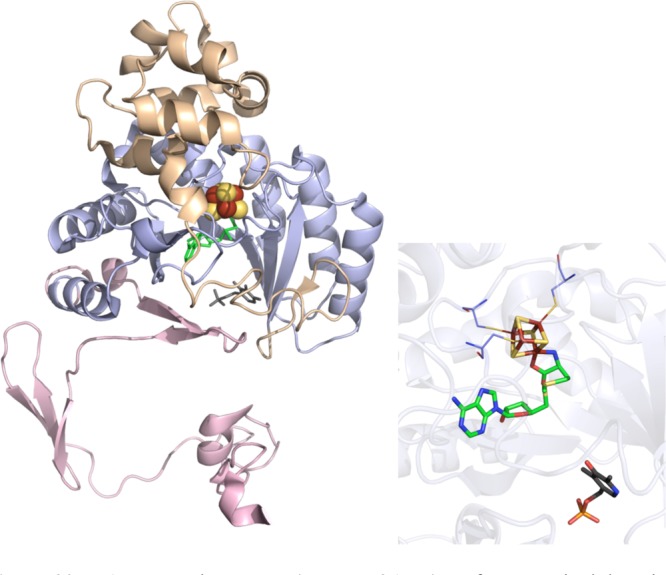
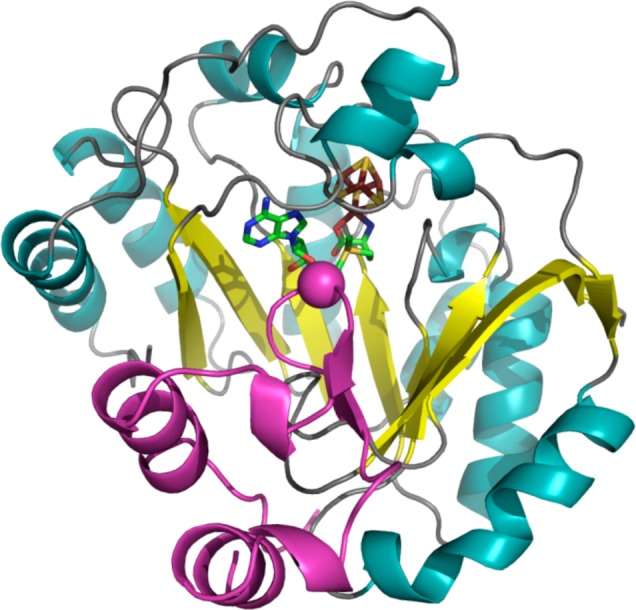
LAM crystal structure
(PDB ID 2A5H). Left: N-terminal domain colored in
wheat, radical SAM domain in light blue, C-terminal domain in light
pink, [4Fe–4S] cluster in yellow and rust spheres, SAM in green
sticks, PLP in dark gray sticks. Right: Active site of LAM where [4Fe–4S]
cluster (yellow and rust), SAM (green carbons), and PLP (gray carbons)
are depicted in sticks with oxygens colored red, nitrogens colored
blue, and phosphorus in orange. The cysteine residues (light blue
carbons) involved in ligating cluster are depicted in lines.
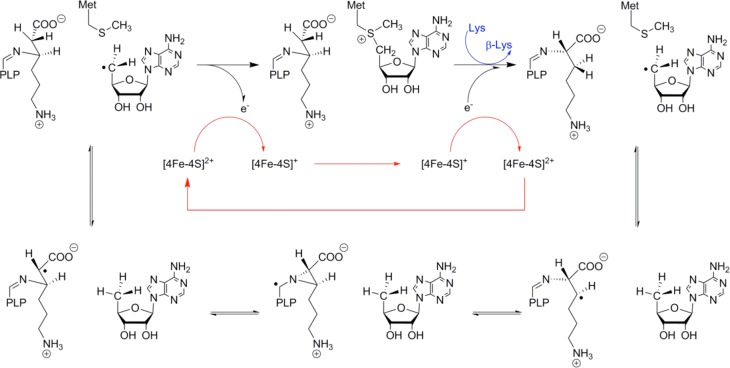
The conversion of l -α-lysine to l -β-lysine
as catalyzed by lysine 2,3-aminomutase.
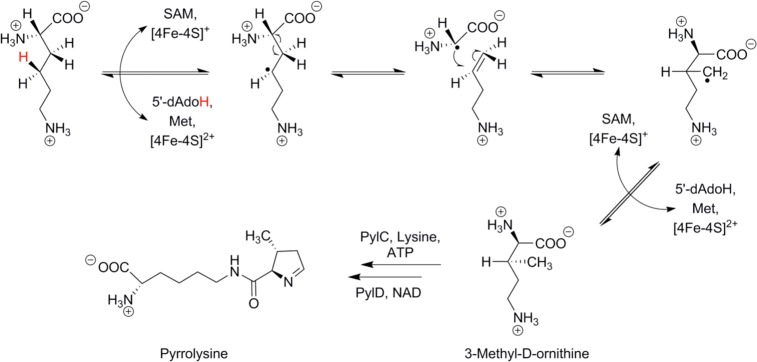
The synthesis of 3-methyl-d -ornithine from l -α-lysine
as catalyzed by PylB in the first step of pyrrolysine biosynthesis.
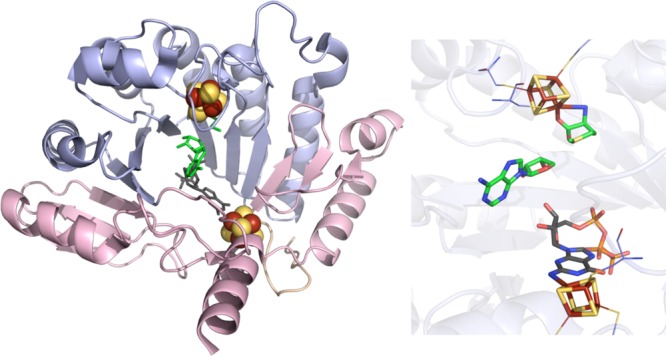
PylB crystal structure
(PDB ID 3T7V). Left: N-terminal domain colored in
wheat, radical SAM domain in light blue, C-terminal domain in light
pink, [4Fe–4S] cluster in yellow and rust spheres, SAM in green
sticks, methylornithine in dark gray sticks. Right: Active site of
PylB where [4Fe–4S] cluster (yellow and rust), SAM (green carbons),
and methylornithine (gray carbons) are depicted in sticks with oxygens
colored red and nitrogens colored blue. Cysteines (light blue carbons)
involved in ligating cluster are depicted in lines.
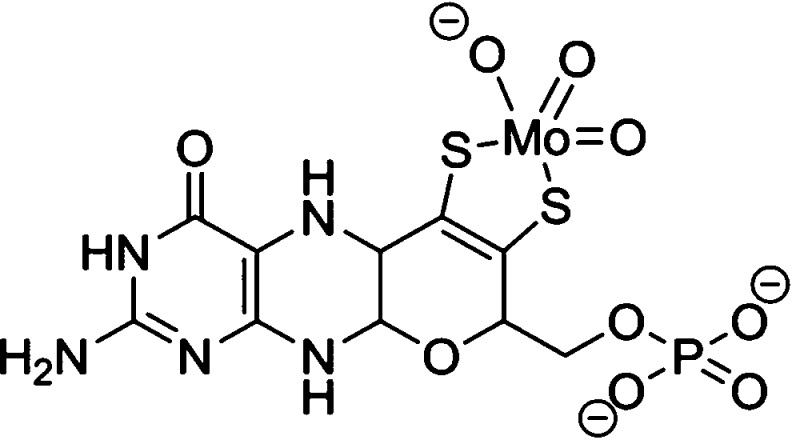
The
molybdopterin cofactor (Moco) is composed of a molybdenum ion
coordinated by a low molecular weight tricyclic pterin ligand via
dithiolene coordination.
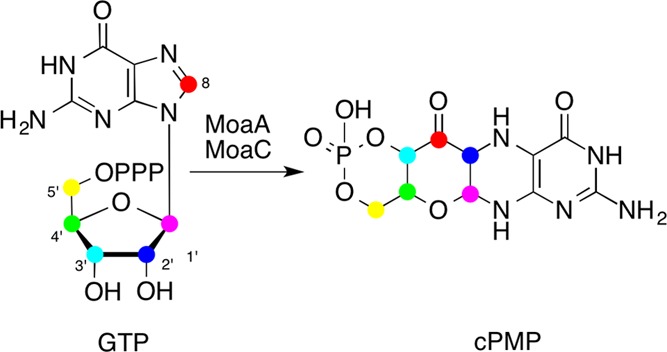
The observed rearrangement of carbon
atoms in the MoaA/MoaC reaction.
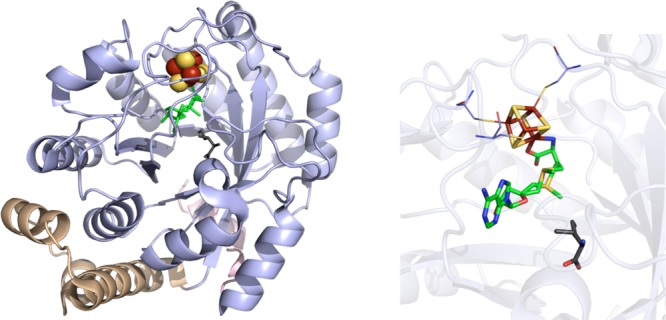
MoaA crystal structure
(PDB ID 2FB3). Left: N-terminal domain colored in
wheat, radical SAM domain in light blue, C-terminal domain in light
pink, [4Fe–4S] clusters in yellow and rust spheres, dAdoH and
Met in green sticks, GTP in dark gray sticks. Right: Active site of
MoaA where [4Fe–4S] cluster (yellow and rust), dAdoH and Met
(green carbons), and GTP (gray carbons) are depicted in sticks with
oxygens colored red, nitrogens colored blue, and phosphates in orange.
Cysteines (light blue carbons) involved in ligating clusters are depicted
in lines.
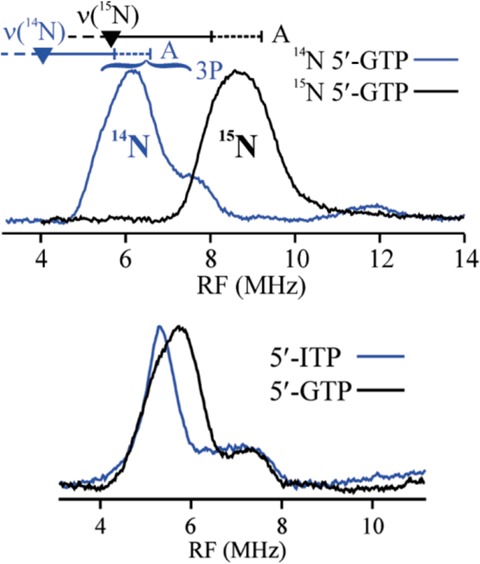
Top
panel: 15N ENDOR evidence of 5′-GTP interaction
with C-terminal [4Fe–4S], using 14N and 15N 5′-GTP substrate and C24S/C28S/C31S MoaA. Bottom panel:
Comparison of 14N ENDOR of substrate analogue 5′-ITP
with 5′-GTP that have equivalent 14N hyperfine interaction
with the C-terminal [4Fe–4S] cluster. Reprinted with permission
from ref (38). Copyright
2006 American Chemical Society.
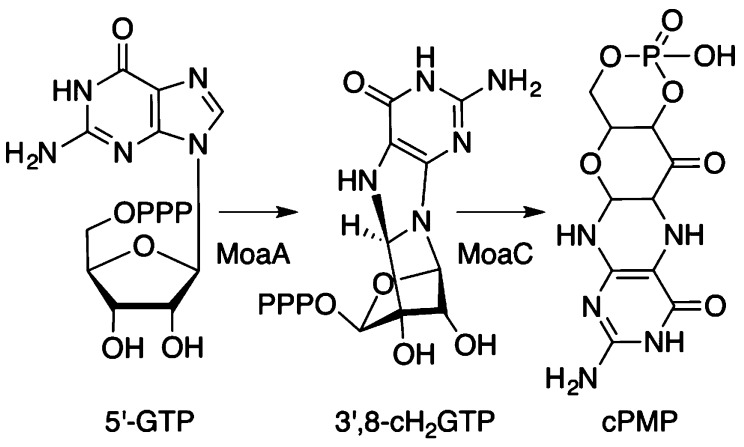
Proposed specific transformations
that are catalyzed by enzymes
MoaA and MoaC.
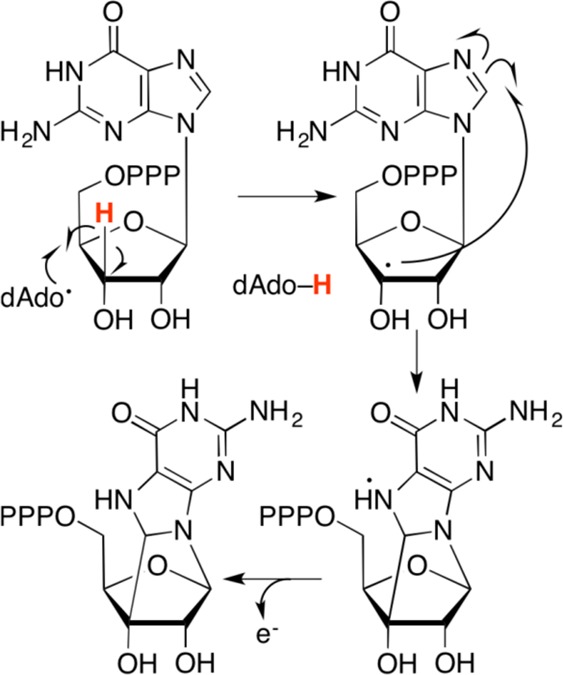
Proposed mechanism of
formation of 3′,8-cH2GTP
by radical SAM enzyme MoaA.
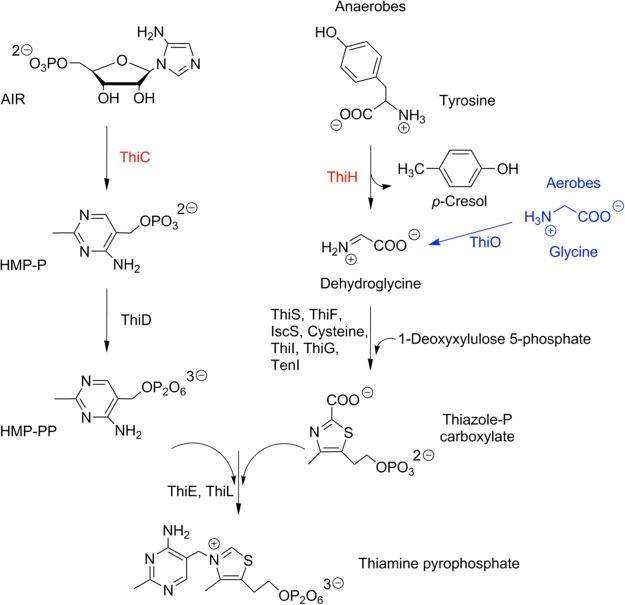
The biosynthesis of thiamine pyrophosphate. 4-Amino-5-hydroxymethyl-2-methylpyrimidine
pyrophosphate (HMP-PP) (left) is ultimately coupled with 4-methyl-5-(β-hydroxyethyl)thiazole
phosphate carboxylate (thiazole-P carboxylate) (right) to form thiamine
pyrophosphate. The radical SAM enzymes ThiC and ThiH are highlighted
in red. The generation of dehydroglycine differs between aerobes and
anaerobes, which is highlighted as well.
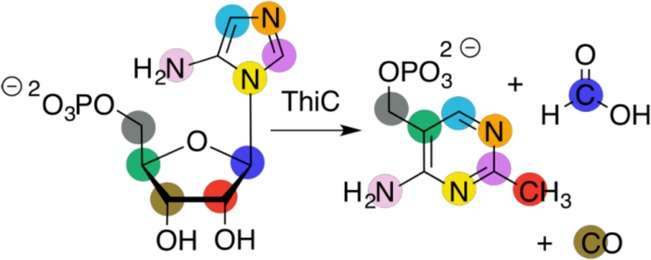
Carbon and nitrogen isotopic label studies in the ThiC
conversion
of AIR to HMP-P. All carbon and nitrogen atoms originate from AIR,
and the two carbon atoms (from the C-1′ and C-3′ positions)
not incorporated into HMP-P produce formic acid and carbon monoxide,
respectively.
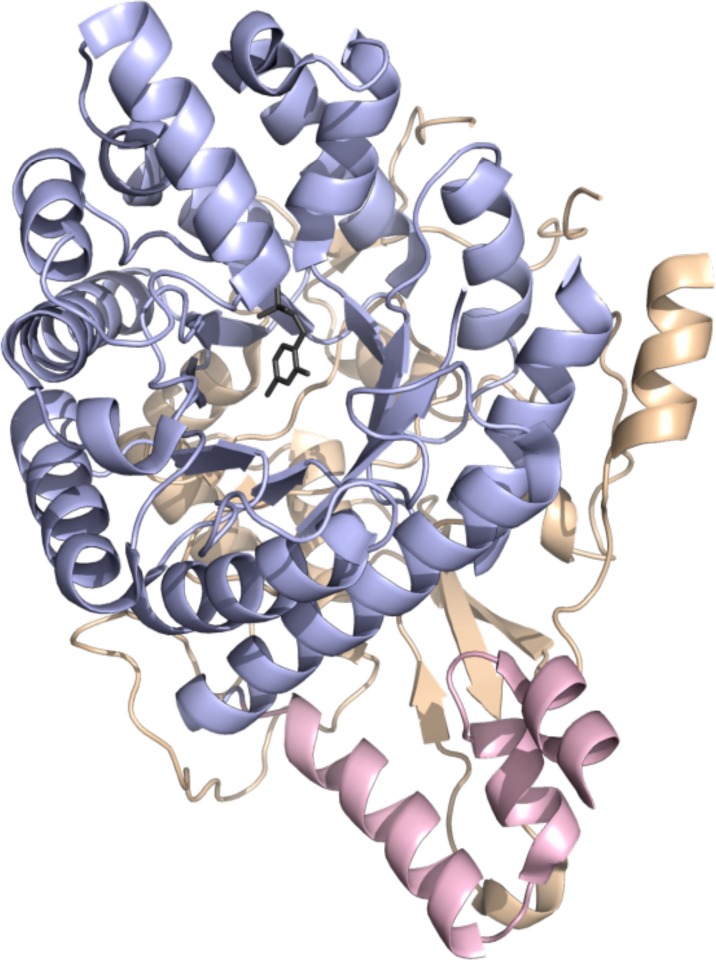
ThiC
crystal structure (PDB ID 3EPO). N-terminal domain colored in wheat,
radical SAM domain in light blue, C-terminal domain in light pink,
HMP-P in dark gray sticks.
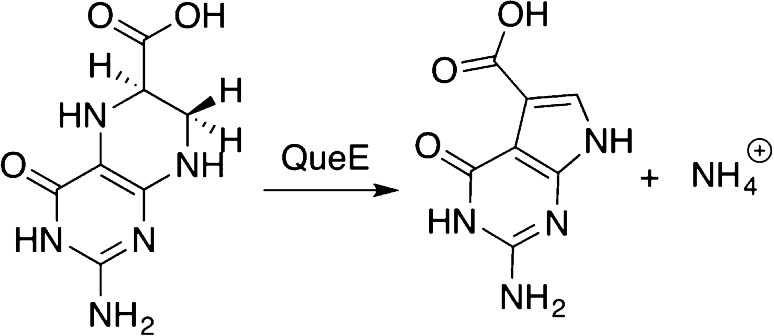
Reaction of the heterocyclic
rearrangement catalyzed by radical
SAM enzyme QueE.
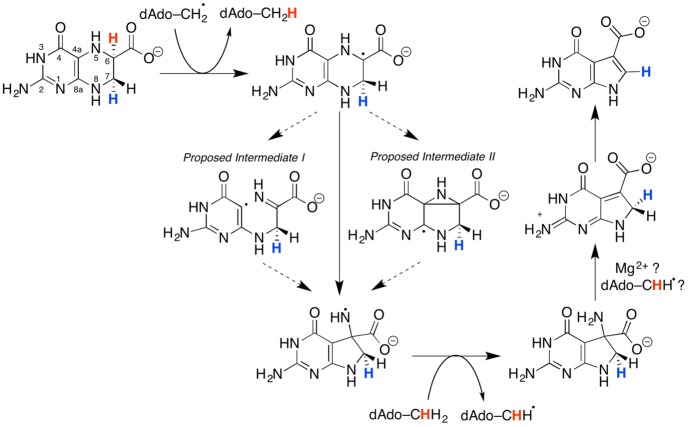
Proposed
mechanism in the QueE heterocyclic rearrangement.
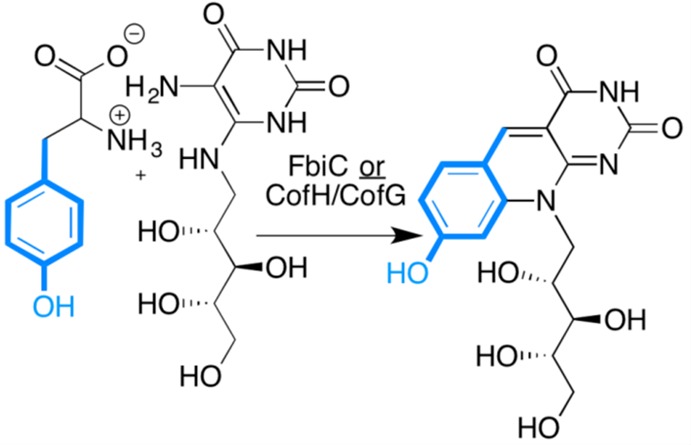
F0 synthase reaction
catalyzed by bifunctional enzyme FbiC. The
monofunctional units of FbiC can be isolated separately and, in vitro,
catalyze the same reaction, with tyrosine and 5-amino-6-ribitylamino-2,4(1H,3H)-pyrimidinedione to make the 8-hydroxy-7-desmethyl-5-deazariboflavin
chromophore F0.
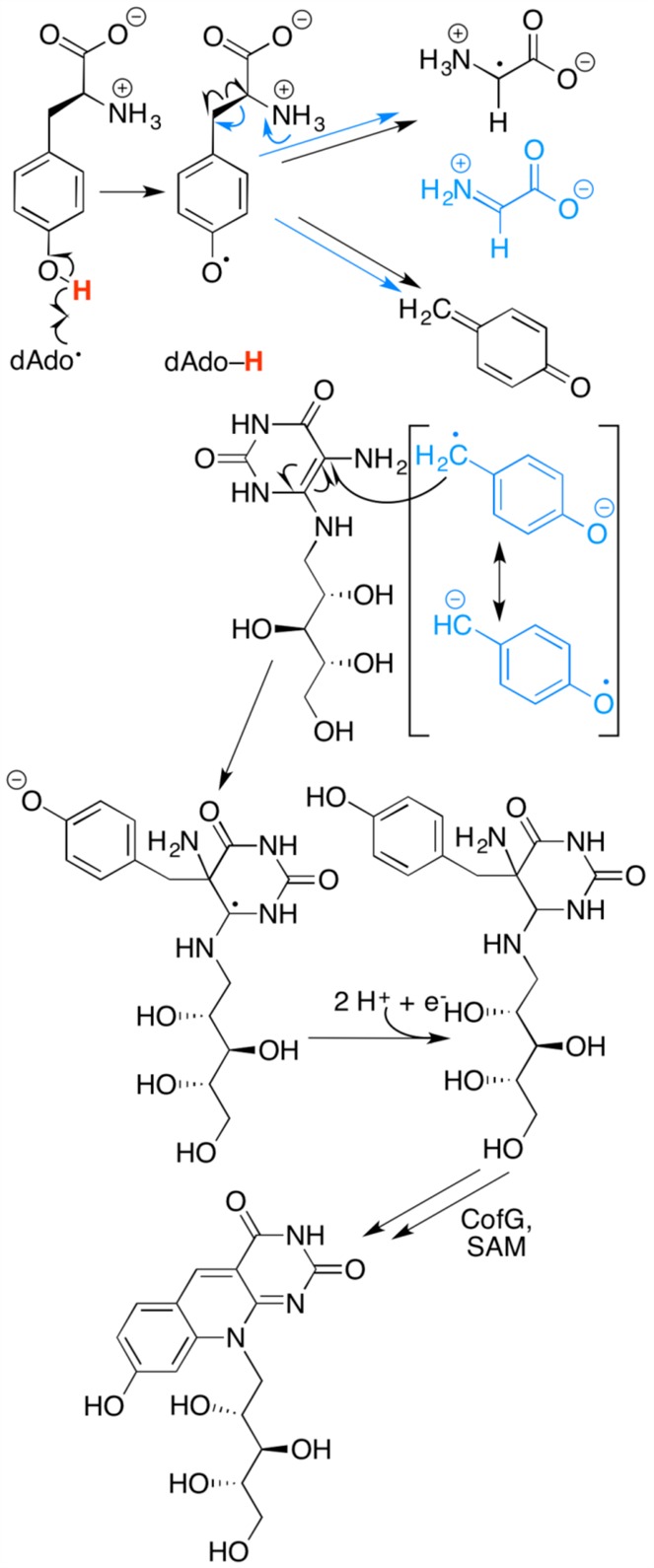
Proposed mechanism of radical initiation and probable
involvement
of CofH. Tyrosine homolytic Cα–Cβ bond cleavage
products are depicted in black, while heterolytic bond cleavage products
are depicted in aqua.
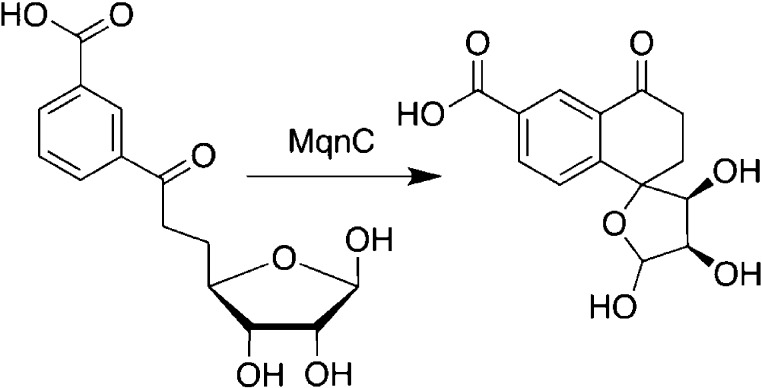
Transformation catalyzed
by radical SAM enzyme MqnC.
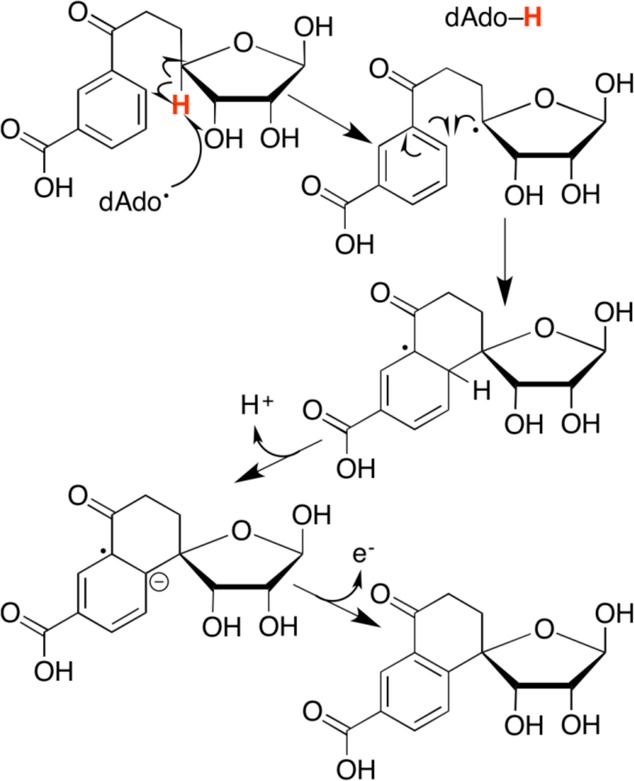
Ring cyclization mechanism catalyzed by radical SAM enzyme MqnC.
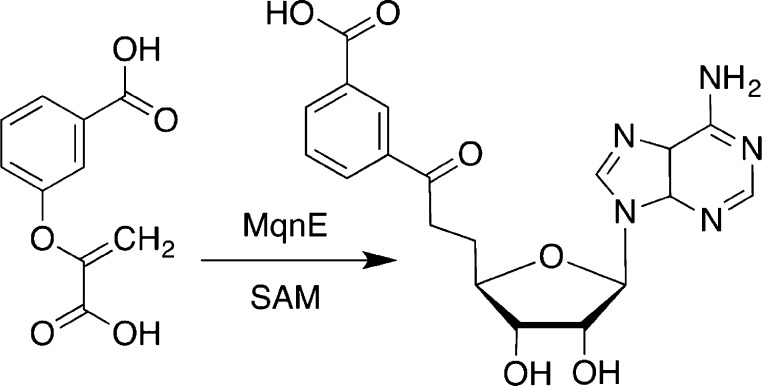
Transformation catalyzed by radical SAM enzyme
MqnE.
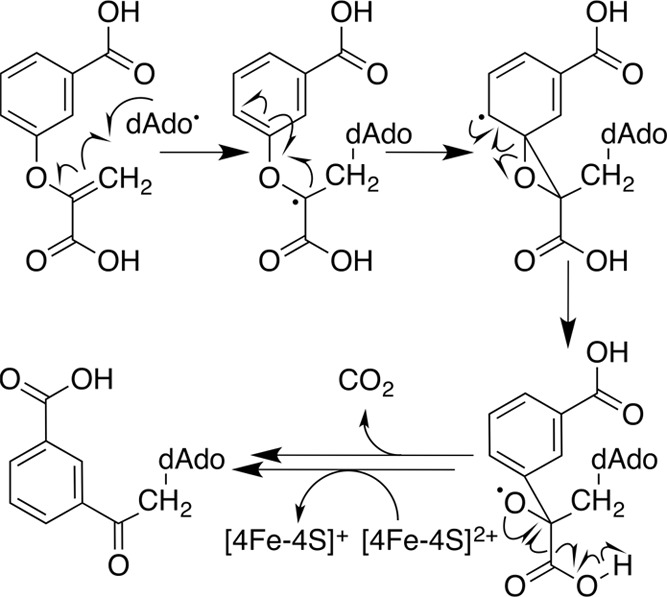
Mechanism of dAdo• addition
to a vinylic ether
double bond, catalyzed by radical SAM enzyme MqnE.

Structure homology model
of the amino acid sequence of MqnE (T. thermophilus) (blue), aligned to the HydE crystal
structure (PDB ID 3CIX) (pink) (section 12.2.5.2). Radical SAM motif
is colored in yellow, cysteines involved in ligating the [4Fe–4S]
are shown as yellow sticks, while the [4Fe–4S] cluster is depicted
as yellow and rust sticks. For clarity, the [2Fe–2S] cluster
of HydE has been omitted. MqnE structural model was generated using
the protein structure prediction server Phyre2, where the HydE template model yielded the top hit.
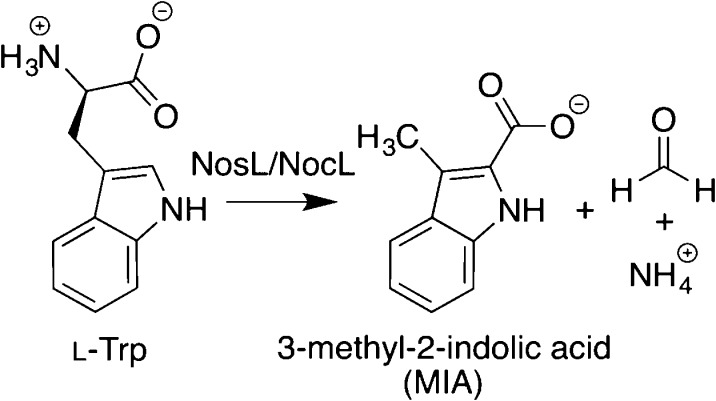
Radical fragmentation–recombination
reaction catalyzed by
radical SAM enzymes NosL and NocL.
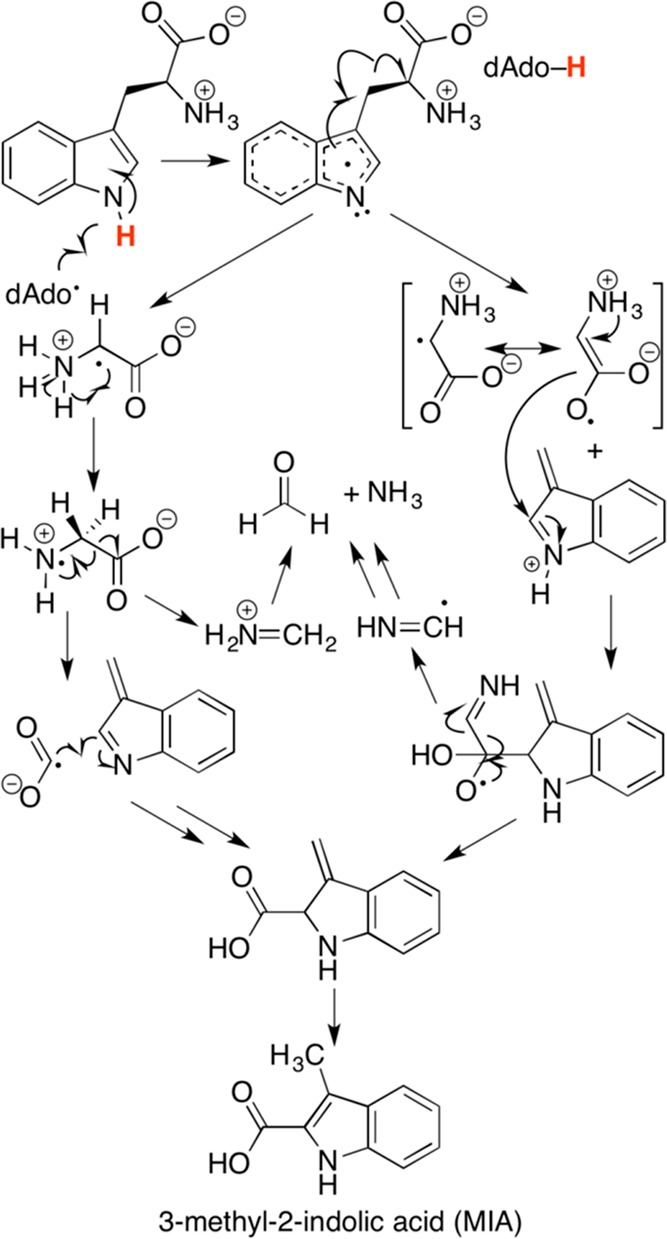
Proposed fragmentation–recombination
mechanism in the conversion
of l -Trp to MIA catalyzed by NosL and NocL.
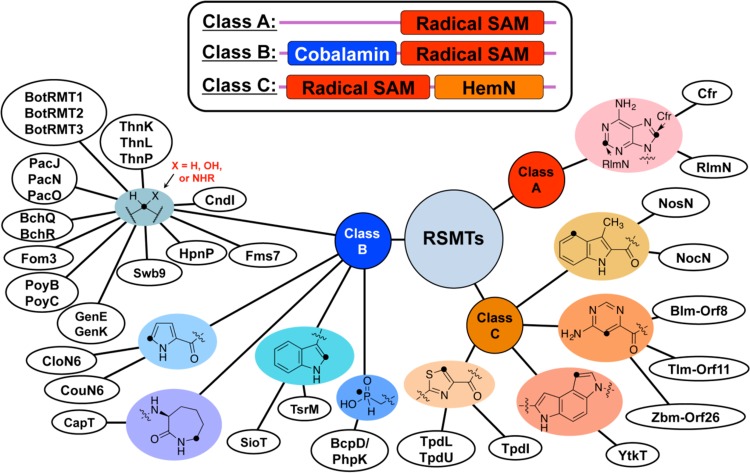
Representative
radical SAM methyltransferase enzymes (RMSTs). Enzymes
are organized by their class, which are differentiated by the top
panel. Members of each class are differentiated by the type of methyl
transfer catalyzed.
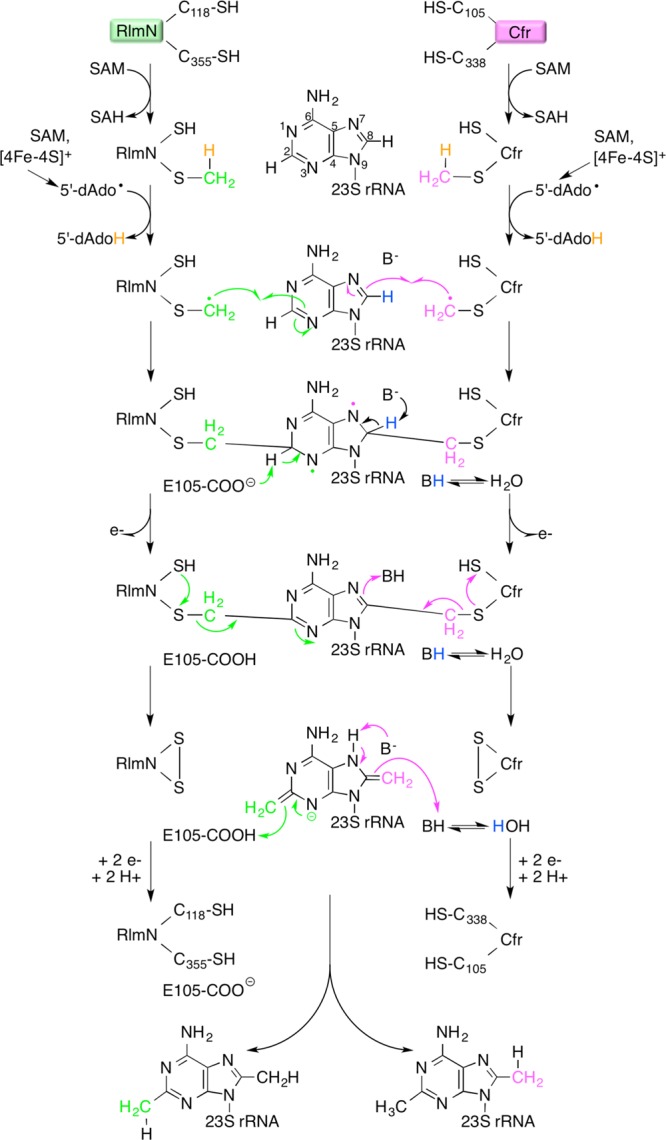
The modification
of adenosine 2503 in 23S RNA of the bacterial
large ribosomal subunit via methylation of the C2 or C8 position as
catalyzed by RlmN or Cfr (two representative members of the class
A RSMTs), respectively. The proposed mechanism for methylation of
C2 by RlmN is depicted along the left side as shown using green arrows.
The proposed mechanism for methylation of C8 by Cfr is depicted along
the right side as shown using magenta arrows.
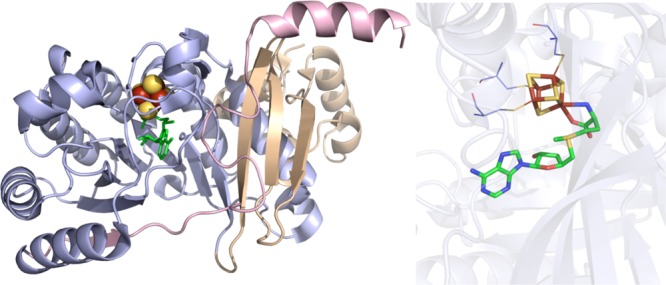
RlmN
crystal structure (PDB ID 3RFA). Left: N-terminal domain colored in
wheat, radical SAM domain in light blue, C-terminal domain in light
pink, [4Fe–4S] cluster in yellow and rust spheres, and SAM
in green sticks. Right: Active site of RlmN where [4Fe–4S]
cluster (yellow and rust) and SAM (green carbons) are depicted in
sticks with oxygens colored red and nitrogens colored blue. Cysteines
(light blue carbons) involved in ligating cluster are depicted in
lines.
Proposed
reaction scheme for class B RSMTs.
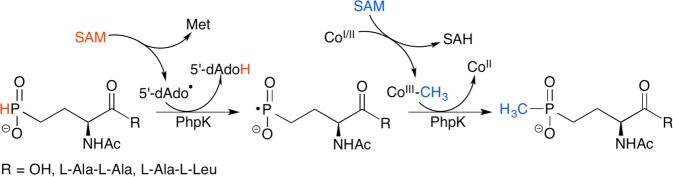
PhpK reaction scheme
catalyzing the conversion of N-acetyldemethylphosphinothricin,
NAcDMPT, to N-acetylphosphinothricin,
NAcPT.
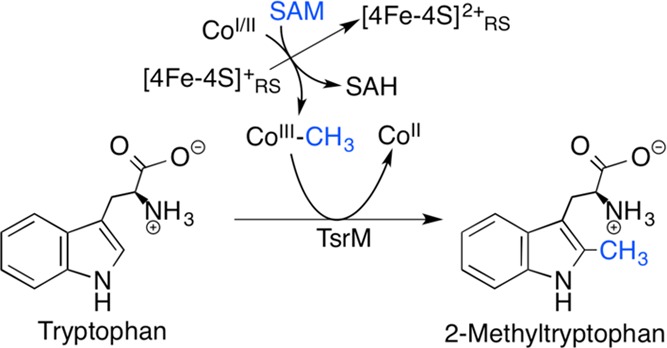
TsrM reaction scheme catalyzing the conversion of tryptophan
to
2-methyltrytophan.
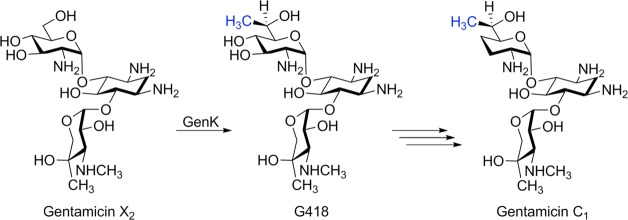
GenK reaction
scheme catalyzing the conversion of Gentamicin X2 to G418.
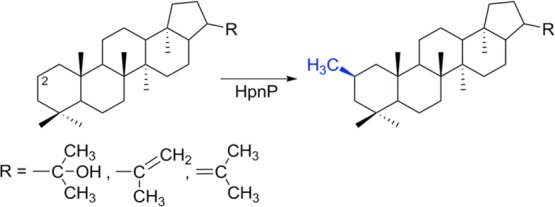
HpnP reaction scheme catalyzing the methylation of the
C2 position
in bacterial hopanoids. A few representative hopanoids are shown here.
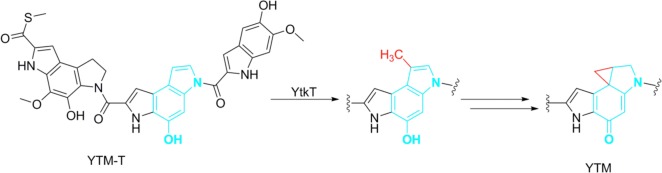
YtkT reaction scheme
catalyzing the methylation of a yatakemycin
(YTM) intermediate prior to cyclopropane ring formation.
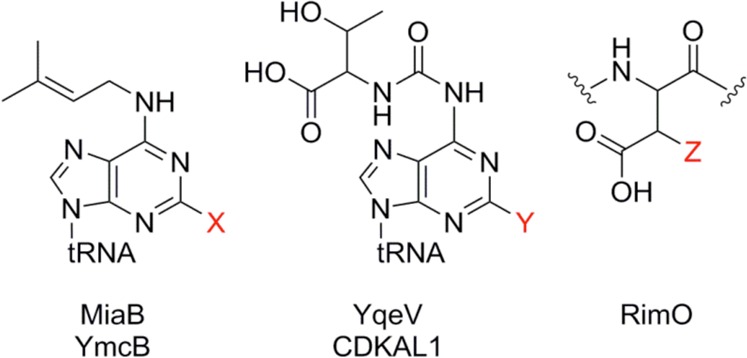
Methylthiolations of nucleic acid and protein residues.
Left: Methylthiolations
as catalyzed by MiaB (E. coli and T. maritima) and YmcB (B. subtilis) where the tRNA adenosine base is modified from i6A (when
X = H) to ms2i6A (when X = SCH3).
Middle: Methylthiolations as catalyzed by YqeV (B.
subtilis) and CDKAL1 (mammalian tRNALys UUU) where the tRNA adenosine base is modified from t6A (when Y = H) to ms2t6A (when Y = SCH3). Right: Methylthiolation as catalyzed by RimO (E. coli ribosomal S12) where Asp89 (when Z = H) is
modified to β-methylthio Asp89 (when Z = SCH3).
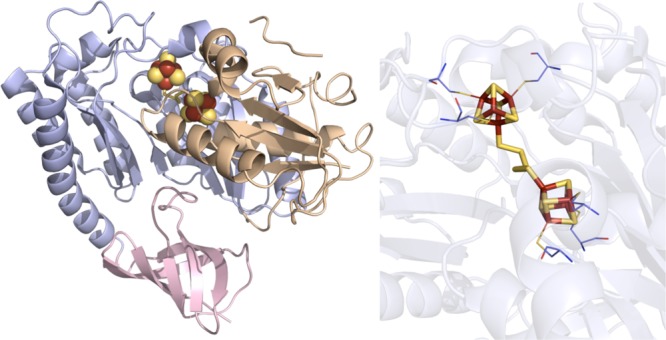
RimO crystal structure (PDB ID 4JC0). Left: N-terminal
UPF0004 domain colored
in light orange, radical SAM domain in light blue, C-terminal TRAM
domain in light pink, [4Fe–4S] clusters in yellow and rust
spheres, and SAM in green sticks. Right: Active site of RimO where
the [4Fe–4S] clusters (yellow and rust) and pentasulfide moiety
(yellow) are depicted in sticks. Cysteines (light blue carbons) involved
in ligating clusters are depicted in lines.
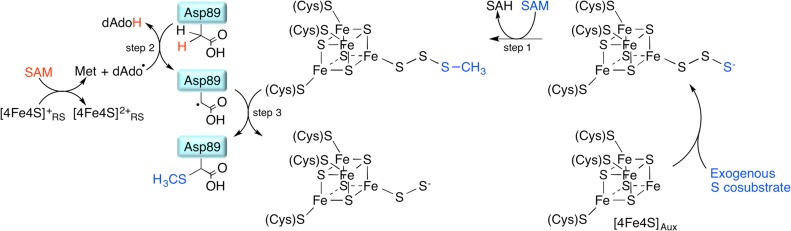
Proposed
RimO mechanism catalyzing the methylthiolation of Asp89
of the ribosomal S12 protein.
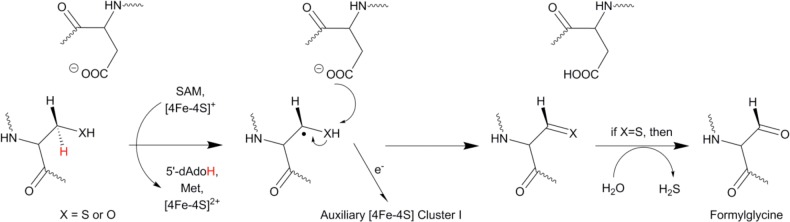
Mechanism
of formylglycine generation for cysteine (anSMEcpe) and
serine type (AtsB/anSMEkp) sulfatase maturating enzymes. The order
of H-atom abstraction and proton abstraction events has not yet been
established.
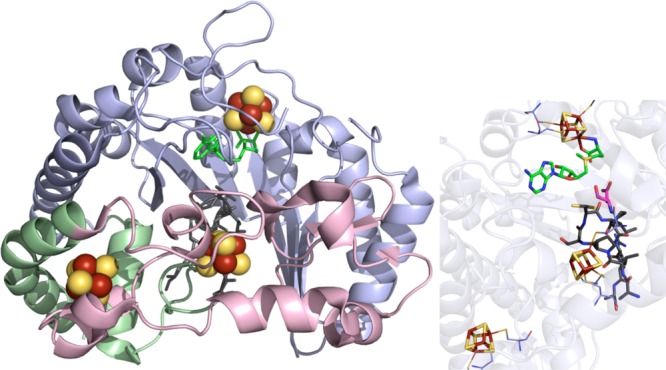
anSME crystal structure (PDB ID 4K39). Left: N-terminal/radical
SAM domain
colored in light blue, SPASM domain in light pink, remaining two α-helices
in light green, [4Fe–4S] clusters in yellow and rust spheres,
SAM in green, and peptide in gray sticks. Right: Active site of anSME
where the [4Fe–4S] clusters (yellow and rust), SAM (green carbons),
and peptide (gray carbons) depicted in sticks with oxygens colored
red and nitrogens colored blue. D277 (magenta carbons, depicted in
sticks) has been identified as the catalytic base (see Figure 66). Cysteines (light blue carbons) involved in ligating
clusters are depicted in lines.
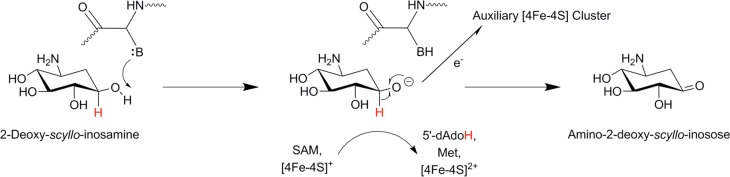
The dehydrogenation reaction catalyzed by BtrN during butirosin
biosynthesis. The order of H-atom abstraction and proton abstraction
events has not yet been definitively established, and along these
lines it has only recently been suggested that formation of the α-hydroxyalkyl
radical by H-atom abstraction may activate the C3-hydroxyl functional
group by decreasing its pKa. Goldman et
al. have identified the putative base involved in catalysis as being
Arg152.
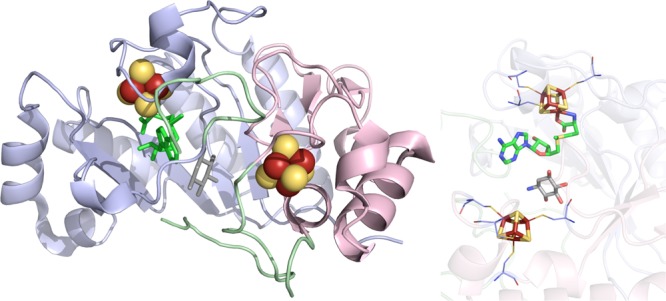
BtrN crystal structure
(PDB ID 4M7T). Left: Radical SAM domain colored in
light blue, C-terminal domain in light pink, linker regions in light
green, [4Fe–4S] clusters in yellow and rust spheres, SAM in
green, and DOIA in gray sticks. Right: Active site of BtrN where the
[4Fe–4S] clusters (yellow and rust), SAM (green carbons), and
DOIA (gray carbons) depicted in sticks with oxygens colored red and
nitrogens colored blue. Cysteines (light blue carbons) involved in
ligating clusters are depicted in lines.
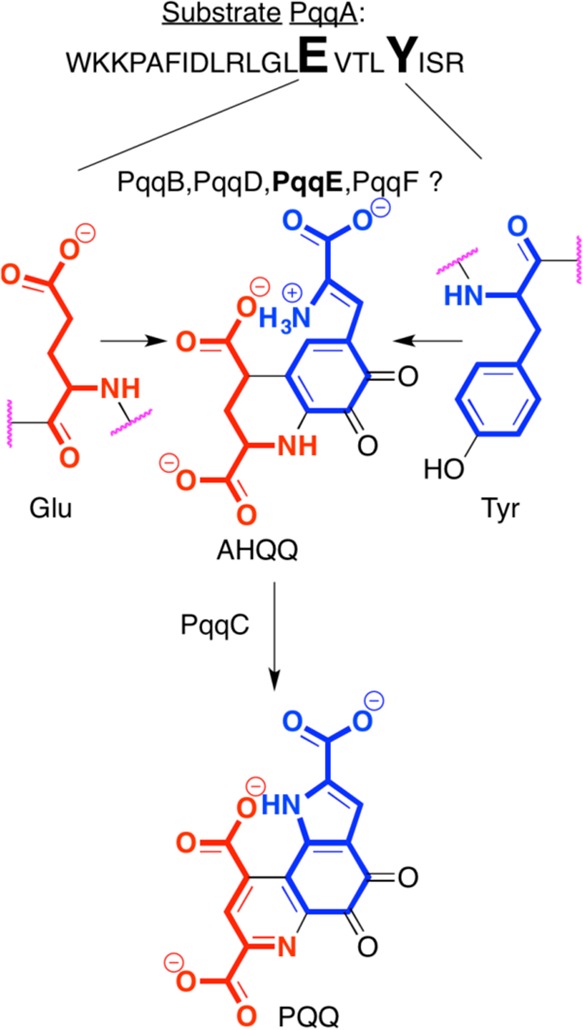
Involvement
of radical SAM enzyme PqqE in pyrroloquinoline quinone
(PQQ) biosynthesis. PqqE is proposed to be involved in the condensation
of peptide residues Glu and Tyr, but its specific substrate is unknown.
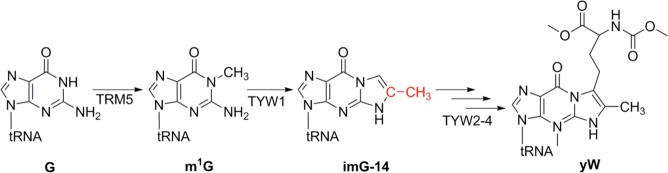
The biosynthesis of yW. G at position 37 is transformed to imG-14
through the actions of TRM5 and TYW1, which is subsequently converted
to yW through the actions of TYW2, TYW3, and TYW4.
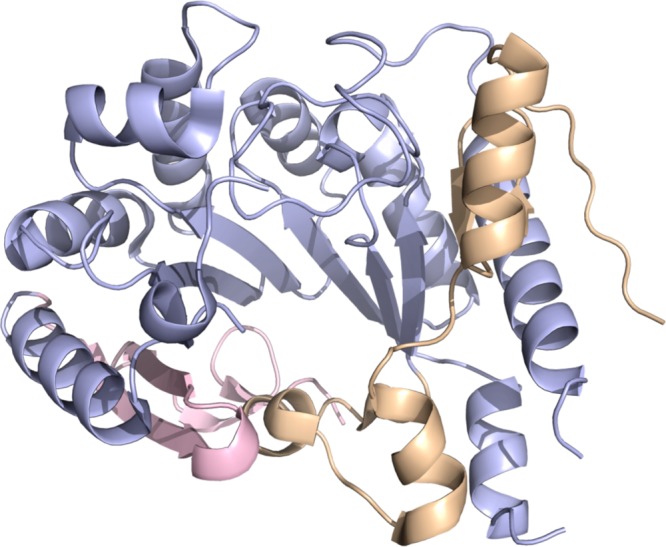
phTYW1 crystal structure (PDB ID 2YX0). N-terminal domain
colored in wheat,
radical SAM domain in light blue, and C-term domain in light pink.
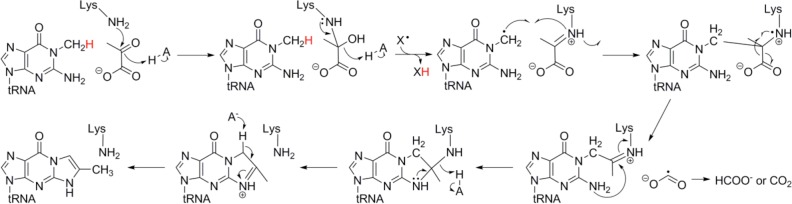
Proposed mechanism for the conversion
of N-methylguanosine
(m1G) to 4-demethylwyosine (img-14) catalyzed by TYW1.
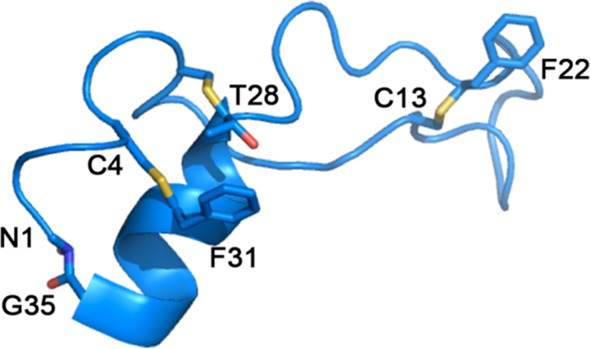
Solution NMR structure of Subtilosin A (PDB ID 1PXQ).
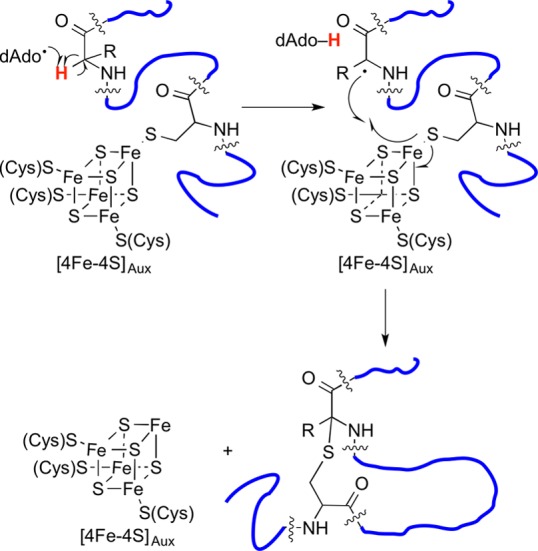
Proposed hydrogen atom abstraction mechanism in thioether
bond
formation observed for radical SAM enzymes AlbA and SkfB.
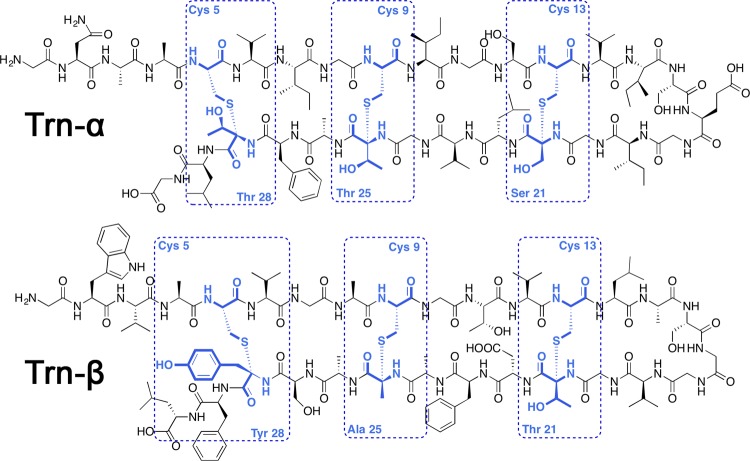
Thuricin CD, like subtilosin A, contains three
thioether cross-links
between cysteine residues and the α-carbons of the modified
amino acids, which are two threonines and a serine in Trnα and
one tyrosine, one alanine, and one threonine in Trnβ.
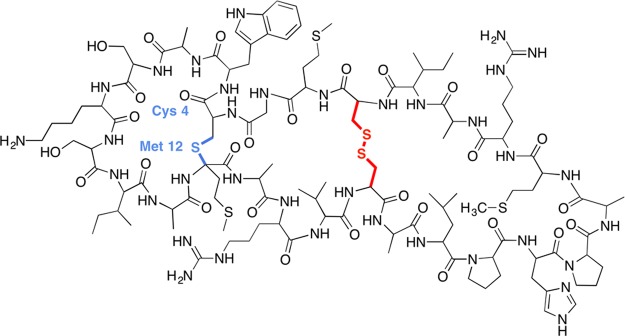
Structure of sporulation killing factor (SKF). The thioether
bond
is highlighted in blue, while the cysteine disulfide bond is highlighted
in red.
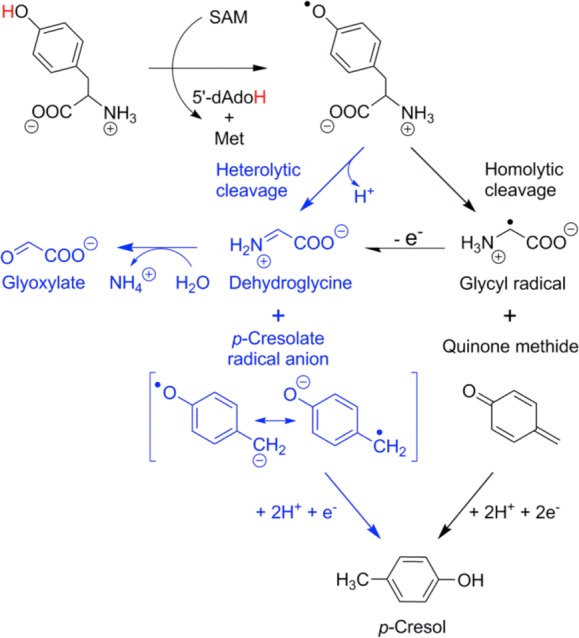
The mechanism of Cα–Cβ
tyrosine bond cleavage
as catalyzed by ThiH. Bond breakage may either occur through a heterolytic
process forming dehydroglycine directly or through a homolytic process
forming a glycyl radical.
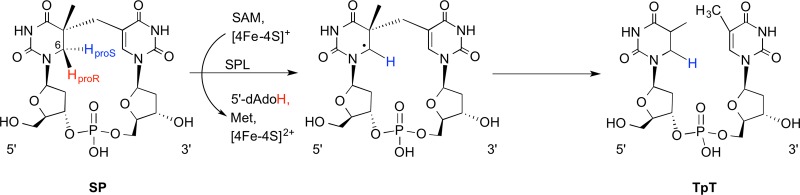
SPL reaction
scheme catalyzing the conversion of SP to TpT.
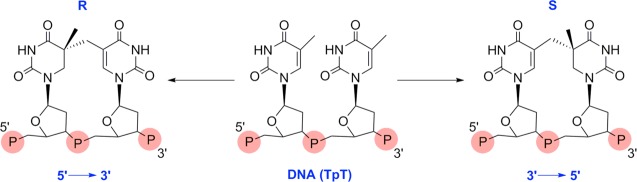
Depiction of the two possible spore photoproducts
with either 5R or 5S configuration.
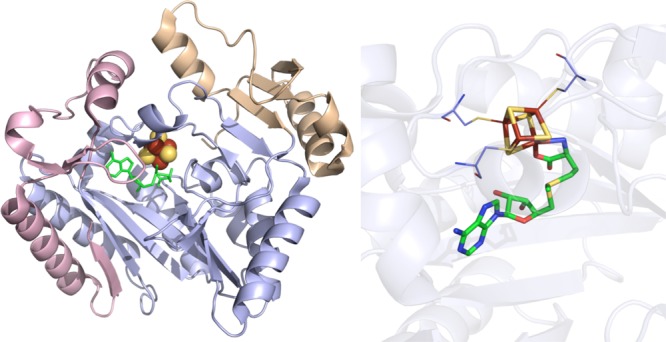
SPL crystal
structure (PDB ID 4FHD). Left: N-terminal domain colored in
wheat, radical SAM domain in light blue, C-terminal domain in light
pink, [4Fe–4S] cluster in yellow and rust spheres, and SAM
in green sticks. Right: Active site of SPL where [4Fe–4S] cluster
(yellow and rust) and SAM (green carbons) are depicted in sticks with
oxygens colored red and nitrogens colored blue. Cysteines (light blue
carbons) involved in ligating cluster are depicted in lines.
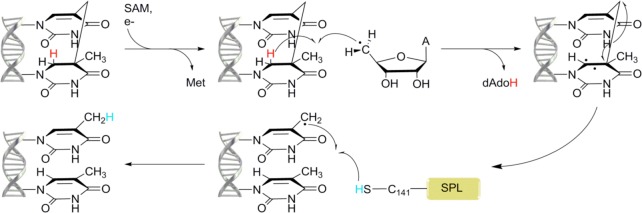
Proposed
mechanism for the repair of SP as catalyzed by SPL.
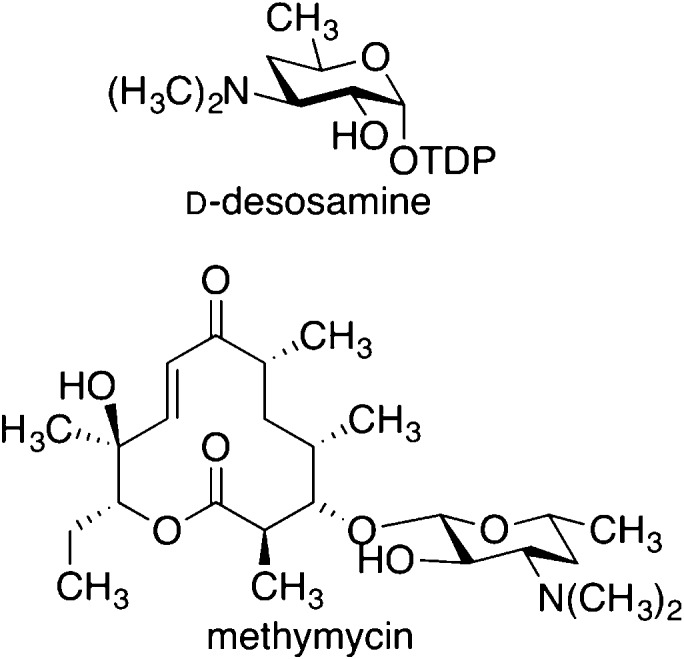
Natural
products d -desosamine and methymycin. Radical
SAM enzyme DesII is involved in the biosynthesis of d -desosamine.
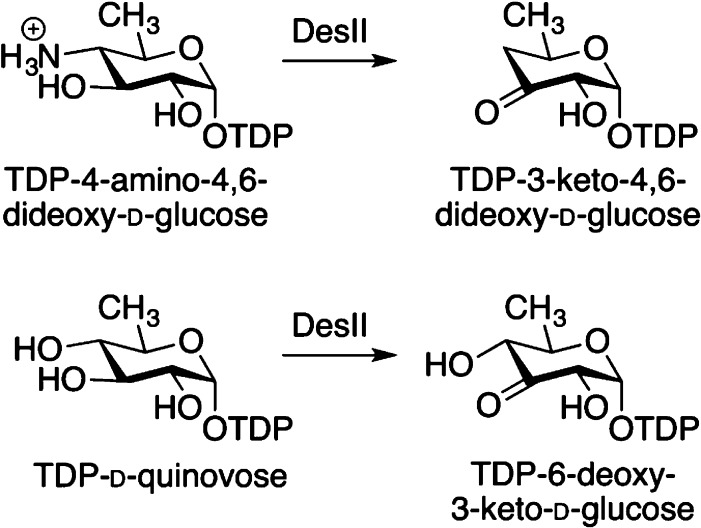
Radical SAM enzyme DesII catalyzes a redox-neutral deamination
(top reaction), as well as an oxidative dehydrogenation (bottom reaction).
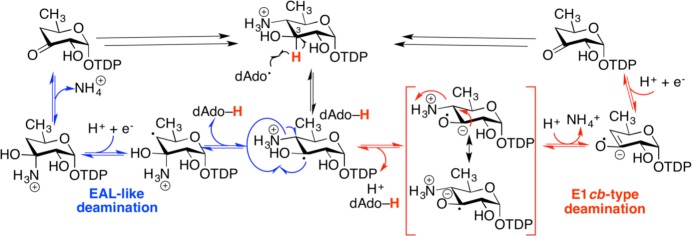
Proposed mechanism in DesII-catalyzed deamination. Depicted in
blue is an ethanolamine ammonia lyase-inspired mechanism involving
the formation of a carbinolamine intermediate. Depicted in red is
an E1cb-type mechanism involving a stabilized enol
radical. Double black full arrows represent product leaving the active
site, substrate coordination, and dAdo• generation.
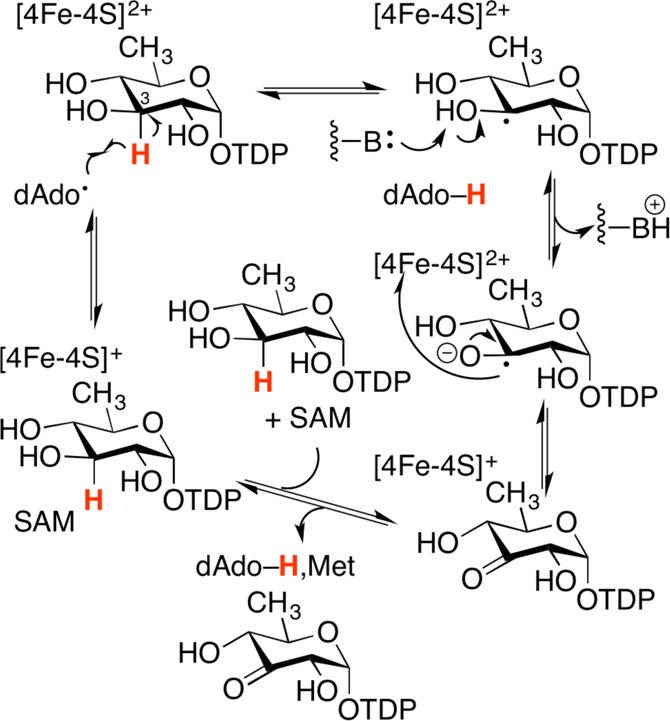
Proposed mechanism of DesII-catalyzed
oxidative dehydrogenation.
Note that electron transfer is proposed to occur from the product
radical back to the oxidized [4Fe–4S]2+ cluster,
as redox cycling of the FeS cluster has been demonstrated.
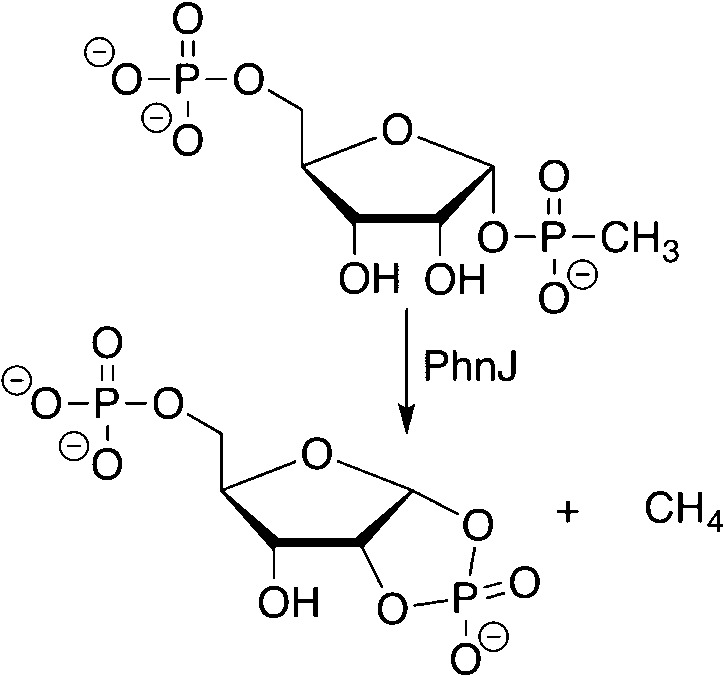
C–P
bond cleavage of methylphosphonate by radical SAM enzyme
PhnJ.
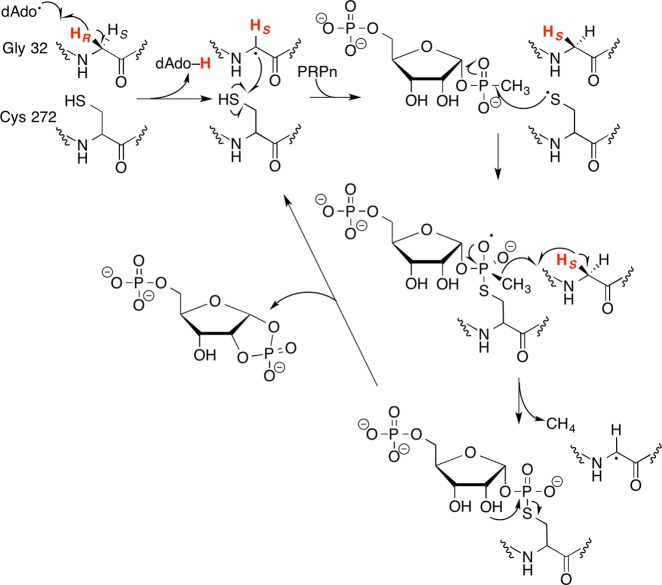
Mechanism of C–P bond cleavage
catalyzed by radical SAM
enzyme PhnJ.
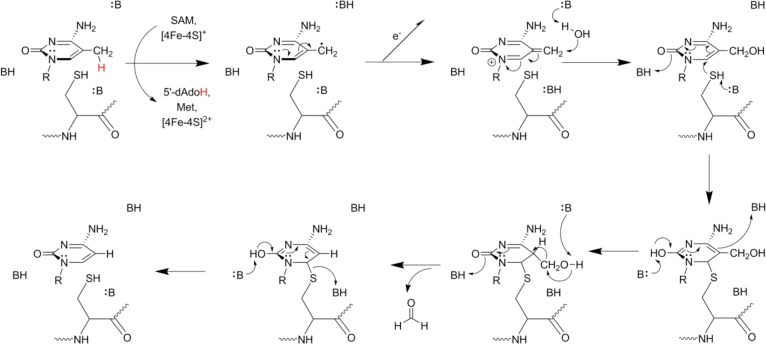
Hypothetical mechanism for the Elp3-catalyzed
demethylation of
DNA. An alternative pathway has been proposed, which could account
for product formation in the absence of an external nucleophile; see
Wu and Zhang.
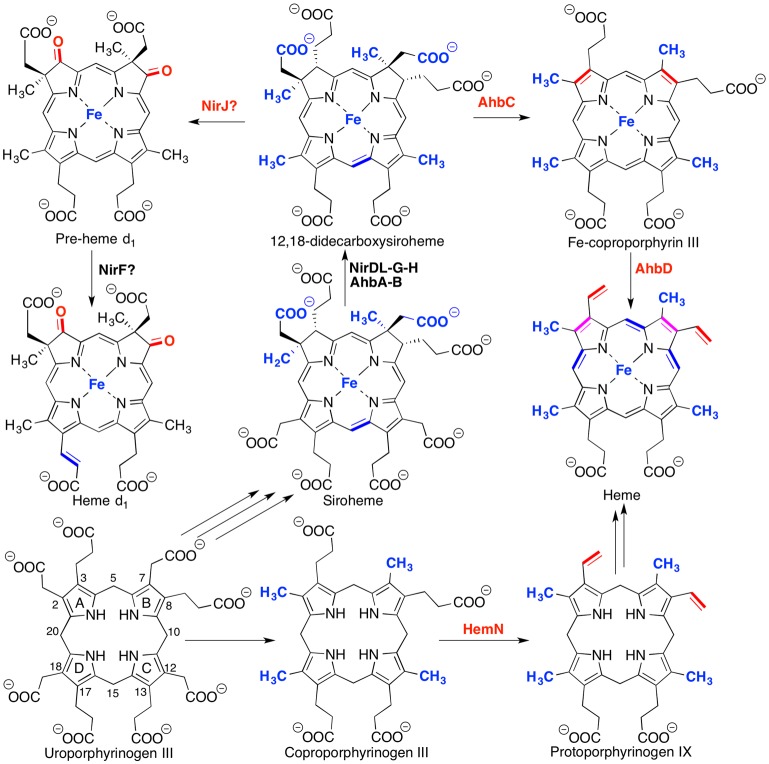
Oxygen-independent biosynthetic
pathway of heme and heme d1 from uroproporphyrinogen III.
Biosynthetic involvement of
radical SAM enzymes and the transformation catalyzed is bolded in
red, while nonradical SAM enzyme transformations are bolded in blue.
Pathway-dependent transformations that do not strictly require radical
SAM enzyme involvement are highlighted in purple.
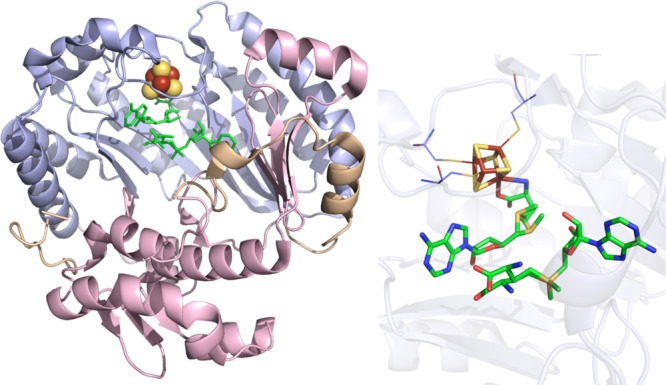
HemN crystal structure (PDB ID 1OLT). Left: N-terminal domain colored in
wheat, radical SAM domain in light blue, C-terminal domain in light
pink, [4Fe–4S] cluster in yellow and rust spheres, and SAM
in green sticks. Right: Active site of HemN where the [4Fe–4S]
cluster (yellow and rust) and SAM (green carbons) are depicted in
sticks with oxygens colored red and nitrogens colored blue. Cysteines
(light blue carbons) involved in ligating clusters are depicted in
lines.
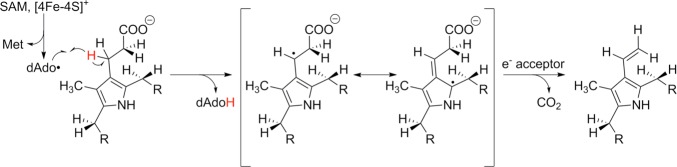
The mechanism of iterative decarboxylation events by HemN.
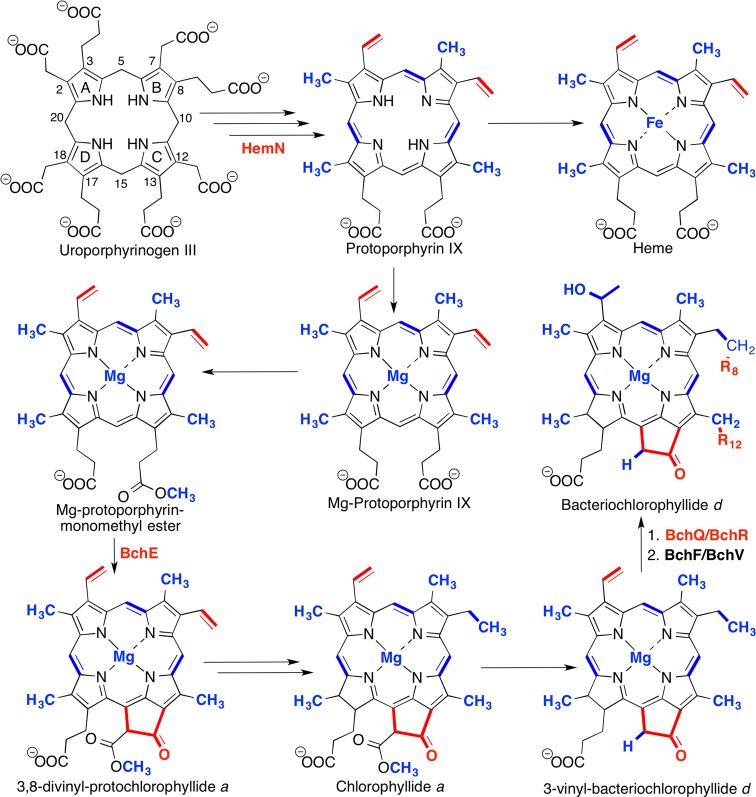
Chlorophyll and bacteriochlorophyll
biosynthetic pathway. Involvement
of radical SAM enzymes (and their respective transformation catalyzed)
is bolded in red, while nonradical SAM enzyme involvement is bolded
in blue.
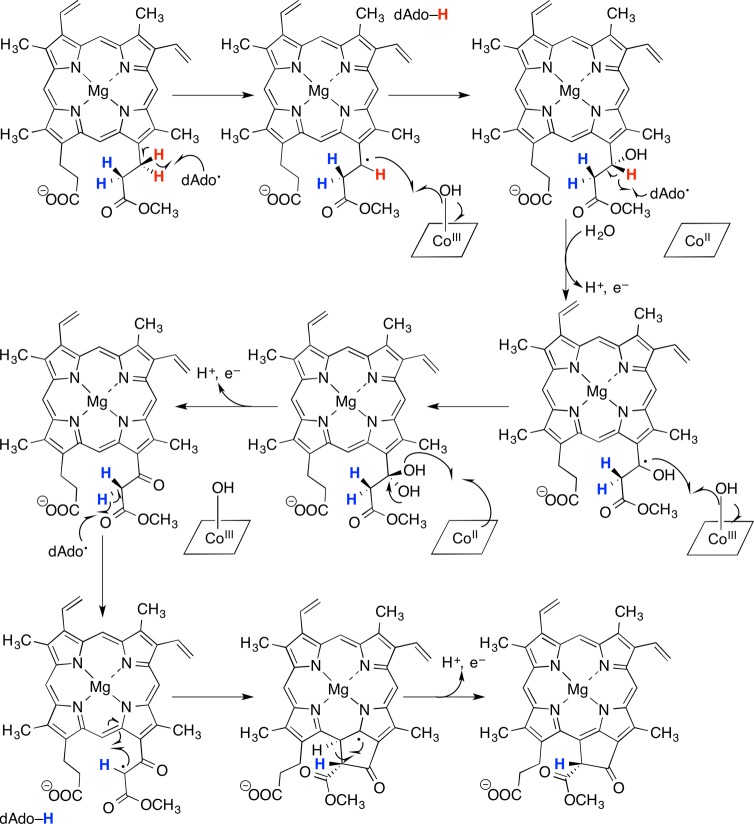
Putative mechanism of
BchE-catalyzed ring cyclization, where both
the radical SAM Fe–S cluster and the cobalamin-binding domains
participate in the ring cyclization reaction.
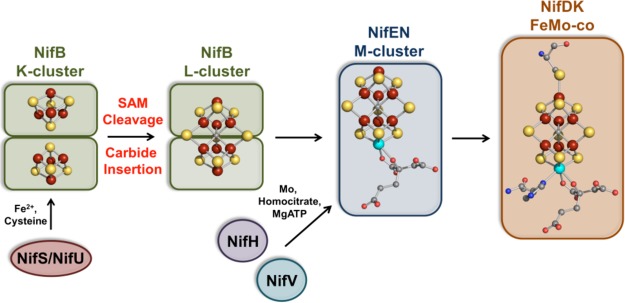
Nitrogenase FeMo-cofactor
assembly.
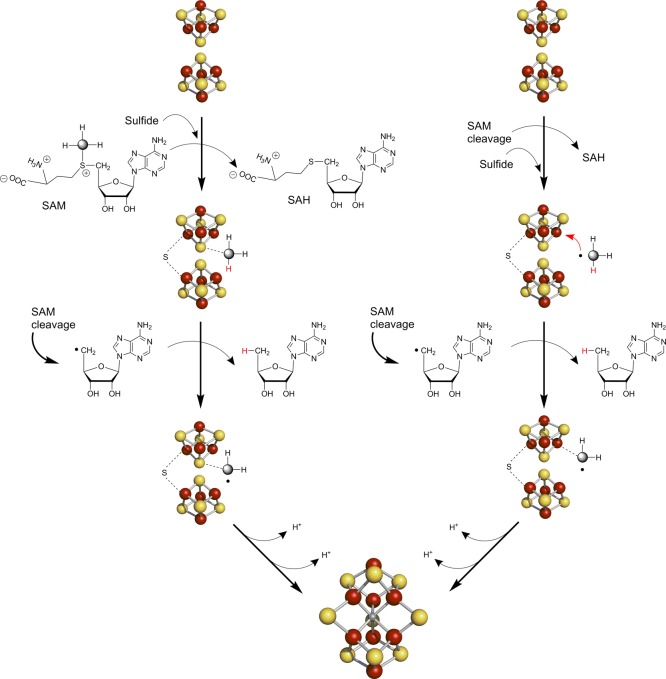
Radical SAM-based carbide insertion during FeMo-co biosynthesis.
NifB is proposed to form the [Fe8–S9]
L-cluster from the K-cluster, two [4Fe–4S] precursor units.
The left pathway invokes methylation of a cluster sulfide followed
by generation of a methylene radical upon hydrogen atom abstraction
by the dAdo•. The right pathway proposes methyl
radical formation via SAM cleavage followed by addition to an Fe ion
of the cluster where further processing to a methylene radical occurs.
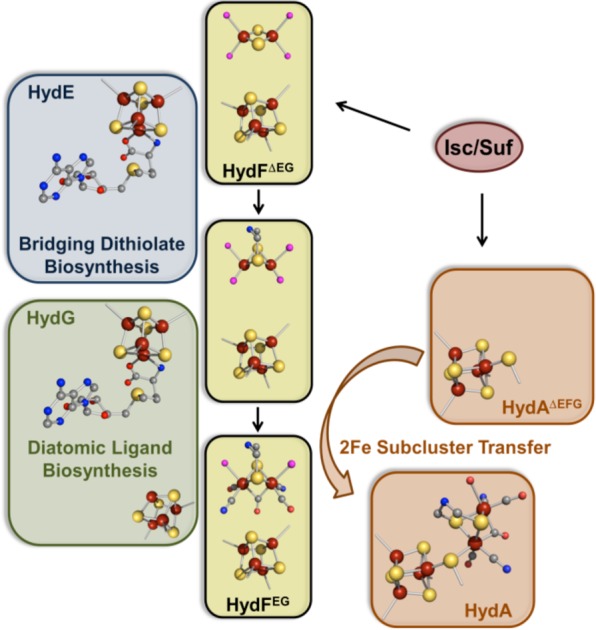
The proposed
maturation pathway for the biosynthesis of the [FeFe]-hydrogenase
H-cluster.
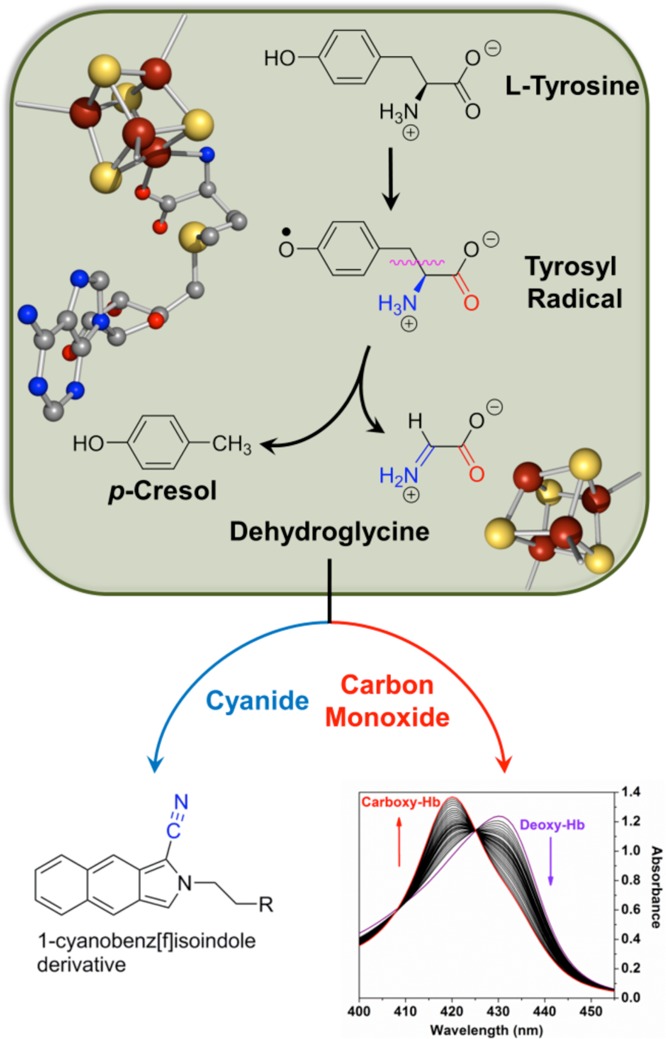
The mechanism of diatomic
ligand biosynthesis as catalyzed by HydG.
Cα–Cβ tyrosine bond cleavage has recently been
demonstrated to occur through a heterolytic process (see text)., Formation of the diatomics requires in some capacity the presence
of the C-terminal [4Fe–4S] cluster where dehydroglycine is
further processed into CN– and CO; the in vitro
complexation/trapping of these diatomics are illustrated at the bottom.
Bottom right reprinted with permission from ref (78b). Copyright 2010 American
Chemical Society.
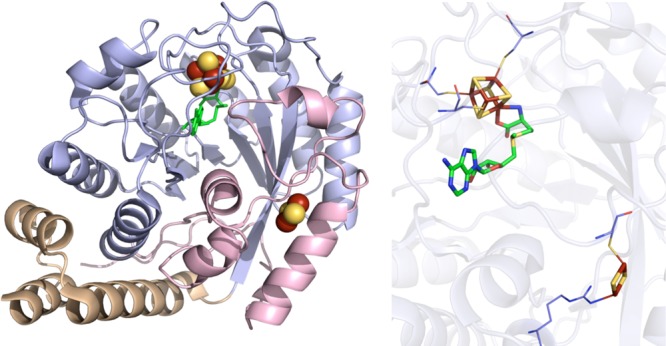
HydE crystal structure (PDB ID 3IIZ). Left: N-terminal domain colored in
wheat, radical SAM domain in light blue, C-terminal domain in light
pink, [4Fe–4S] and [2Fe–2S] clusters in yellow and rust
spheres, and SAM in green sticks. Right: Active site of HydE where
the [4Fe–4S] and [2Fe–2S] clusters (yellow and rust)
and SAM (green carbons) are depicted in sticks with oxygens colored
red and nitrogens colored blue. Cysteines (light blue carbons) involved
in ligating clusters are depicted in lines.
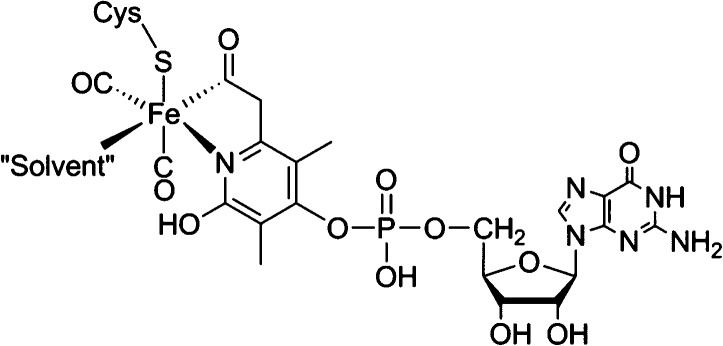
The iron-guanylylpyridinol
cofactor (FeGP-co) of [Fe]-hydrogenase.
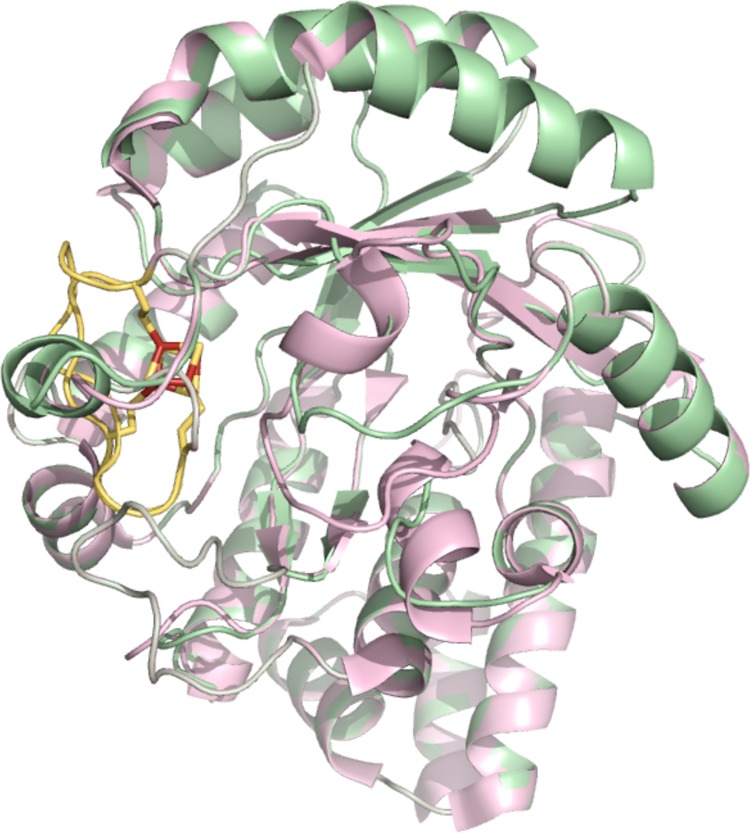
Structure homology
model of the amino acid sequence of HcgA (M. maripaludis) (green), aligned to the HydE crystal
structure (PDB ID 3CIX) (pink) (section 12.2.5.2). Radical SAM motif
is colored in yellow, cysteines involved in ligating the [4Fe–4S]
are shown as yellow sticks, while the [4Fe–4S] cluster is depicted
as yellow and rust sticks. For clarity, the [2Fe–2S] cluster
of HydE has been omitted. MqnE structural model was generated using
the protein structure prediction server Phyre2, where the HydE template model yielded the top hit.

Proposed AviX12 reaction
catalyzing the epimerization reaction
to convert gavibamycin N1 to avilamycin A.
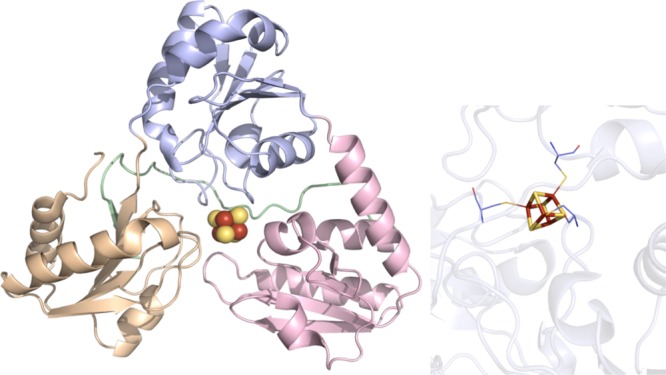
Dph2 crystal
structure (PDB ID 3LZD). Left: Domain 1 colored in wheat, domain
2 in light blue, domain 3 in light pink, C-terminal domain in light
green, and [4Fe–4S] cluster in yellow and rust spheres. Right:
Active site of Dph2 where the [4Fe–4S] cluster (yellow and
rust) is depicted in sticks. Cysteines (light blue carbons) involved
in ligating clusters are depicted in lines.
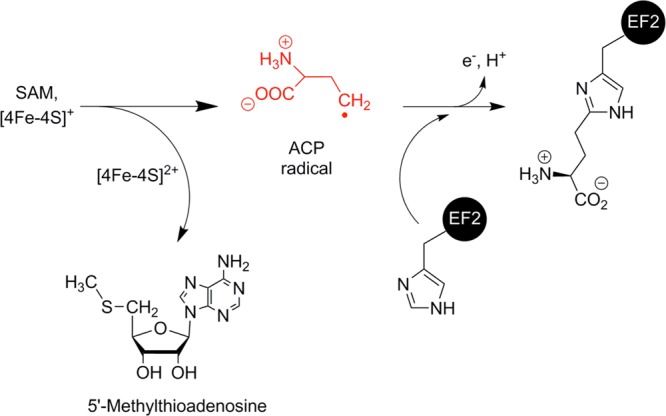
Proposed reaction mechanism
for Dph2 catalyzing the modification
of EF2-His600 during the first step of dipthamide biosynthesis.
Similar articles
-
Radical SAM enzymes and radical enzymology.Biochim Biophys Acta. 2012 Nov;1824(11):1151-3. doi: 10.1016/j.bbapap.2012.07.006. Epub 2012 Jul 22. Biochim Biophys Acta. 2012. PMID: 22850428 No abstract available.
-
Adenosylmethionine-dependent iron-sulfur enzymes: versatile clusters in a radical new role.J Biol Inorg Chem. 2001 Mar;6(3):209-26. doi: 10.1007/s007750100210. J Biol Inorg Chem. 2001. PMID: 11315557 Review.
-
Pyruvate formate-lyase activating enzyme: elucidation of a novel mechanism for glycyl radical formation.Arch Biochem Biophys. 2005 Jan 1;433(1):288-96. doi: 10.1016/j.abb.2004.09.028. Arch Biochem Biophys. 2005. PMID: 15581584 Review.
-
Two Fe-S clusters catalyze sulfur insertion by radical-SAM methylthiotransferases.Nat Chem Biol. 2013 May;9(5):333-8. doi: 10.1038/nchembio.1229. Epub 2013 Mar 31. Nat Chem Biol. 2013. PMID: 23542644 Free PMC article.
-
The methylthiolation reaction mediated by the Radical-SAM enzymes.Biochim Biophys Acta. 2012 Nov;1824(11):1223-30. doi: 10.1016/j.bbapap.2011.11.007. Epub 2011 Dec 7. Biochim Biophys Acta. 2012. PMID: 22178611 Free PMC article. Review.
Cited by
-
Carbon extension in peptidylnucleoside biosynthesis by radical SAM enzymes.Nat Chem Biol. 2016 Nov;12(11):905-907. doi: 10.1038/nchembio.2187. Epub 2016 Sep 19. Nat Chem Biol. 2016. PMID: 27642865 Free PMC article.
-
S-adenosylmethionine tRNA modification: unexpected/unsuspected implications of former/new players.Int J Biol Sci. 2020 Sep 30;16(15):3018-3027. doi: 10.7150/ijbs.49302. eCollection 2020. Int J Biol Sci. 2020. PMID: 33061813 Free PMC article. Review.
-
Biosynthesis of the catalytic H-cluster of [FeFe] hydrogenase: the roles of the Fe-S maturase proteins HydE, HydF, and HydG.Chem Sci. 2020 Sep 22;11(38):10313-10323. doi: 10.1039/d0sc04216a. Chem Sci. 2020. PMID: 34123177 Free PMC article. Review.
-
Trapping a cross-linked lysine-tryptophan radical in the catalytic cycle of the radical SAM enzyme SuiB.Proc Natl Acad Sci U S A. 2021 May 25;118(21):e2101571118. doi: 10.1073/pnas.2101571118. Proc Natl Acad Sci U S A. 2021. PMID: 34001621 Free PMC article.
-
Radical new paradigm for heme degradation in Escherichia coli O157:H7.Proc Natl Acad Sci U S A. 2016 Oct 25;113(43):12138-12143. doi: 10.1073/pnas.1603209113. Epub 2016 Oct 10. Proc Natl Acad Sci U S A. 2016. PMID: 27791000 Free PMC article.
References
-
- Knappe J.; Schacht J.; Mockel W.; Hopner T.; Vetter H. J.; Edenharder R. Eur. J. Biochem. 1969, 11, 316. - PubMed
-
- Broderick J. B.; Duderstadt R. E.; Fernandez D. C.; Wojtuszewski K.; Henshaw T. F.; Johnson M. K. J. Am. Chem. Soc. 1997, 119, 7396.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
