

Silencing motifs in the Clr2 protein from fission yeast, Schizosaccharomyces pombe
- PMID: 24475199
- PMCID: PMC3903592
- DOI: 10.1371/journal.pone.0086948
Silencing motifs in the Clr2 protein from fission yeast, Schizosaccharomyces pombe
Abstract
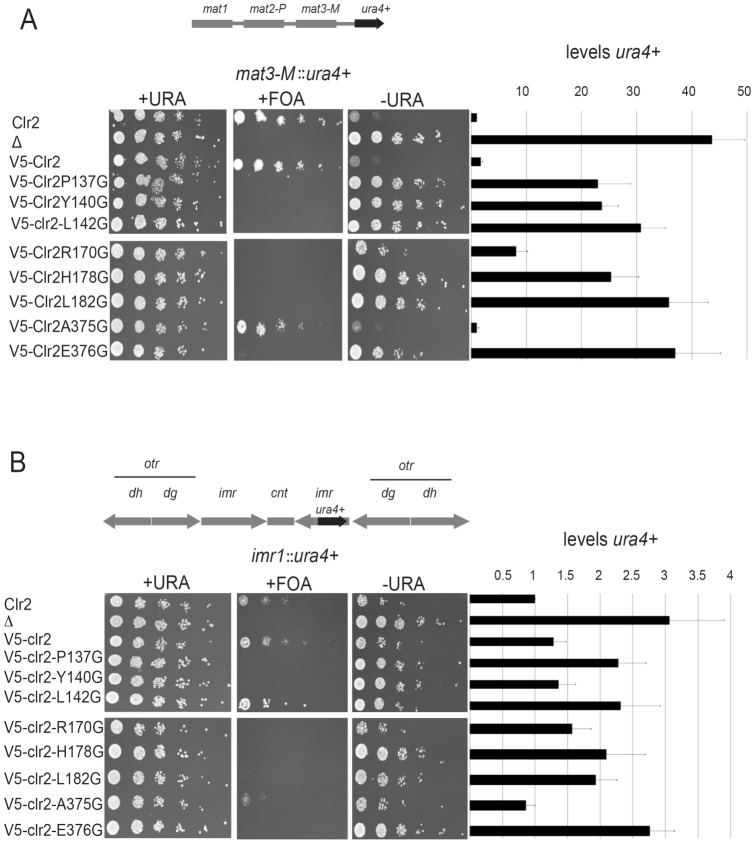
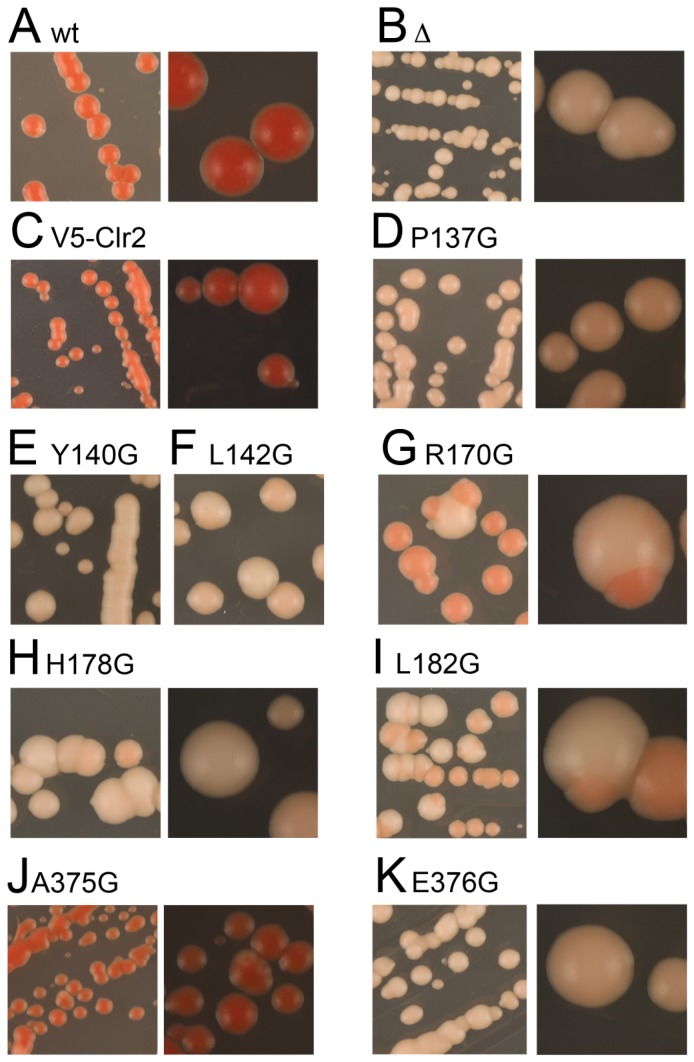
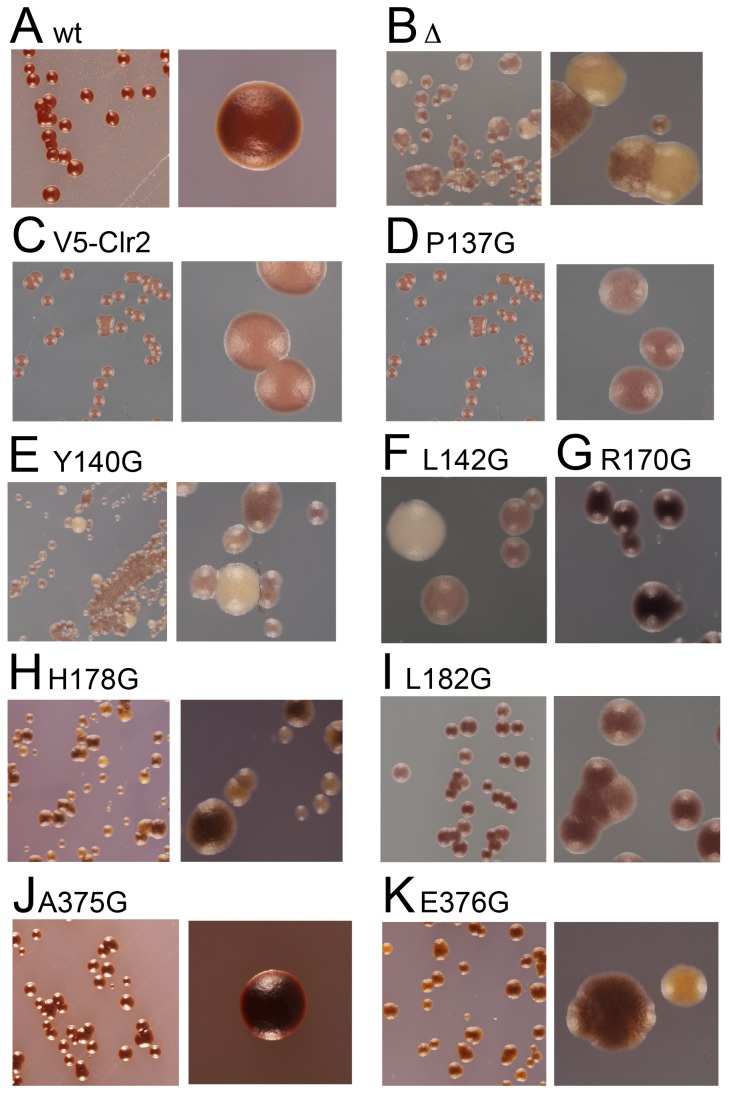
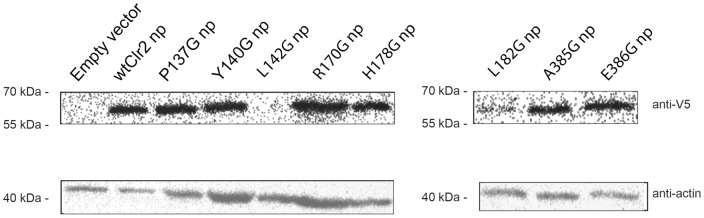
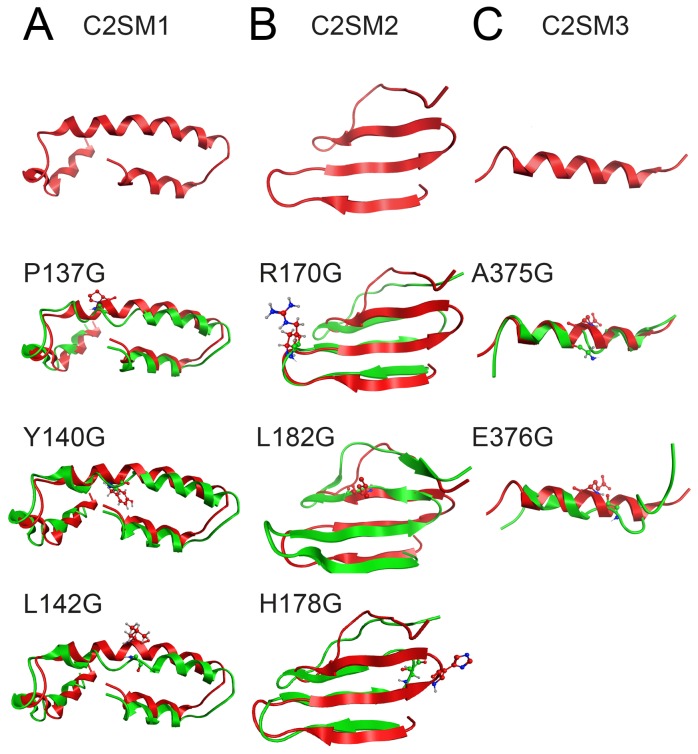
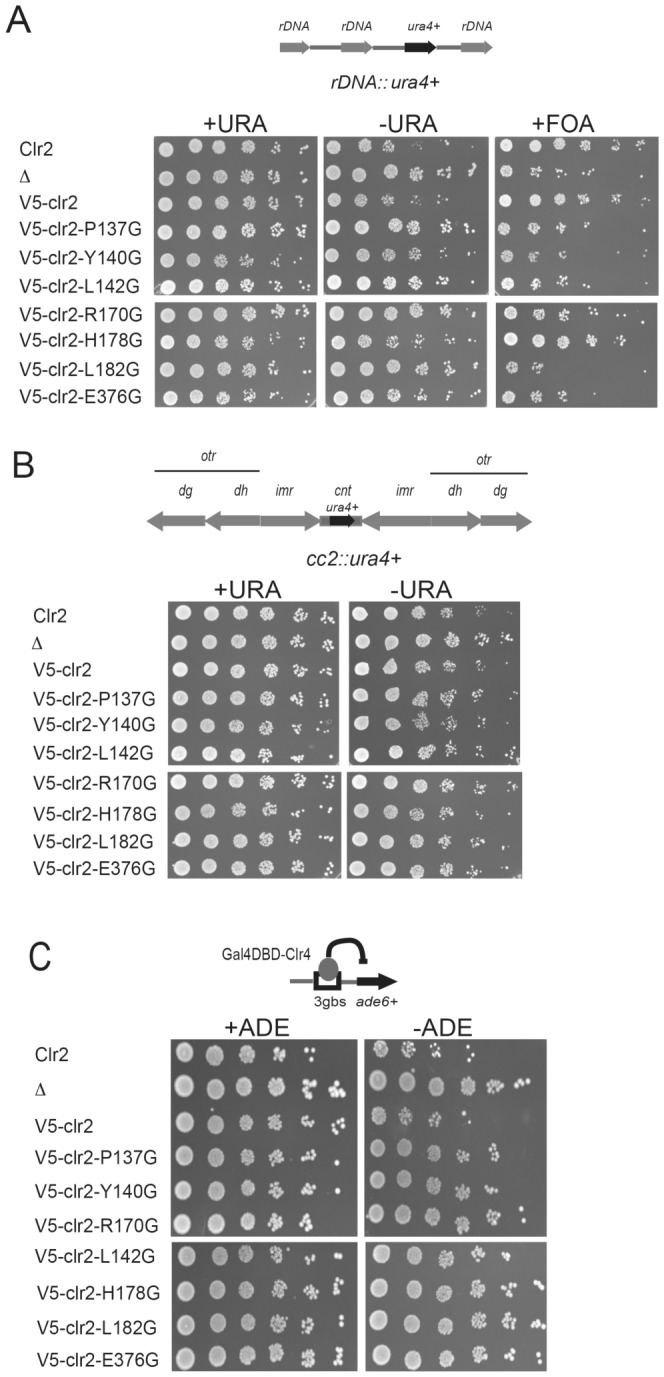
The fission yeast, Schizosaccharomyces pombe, is a well-established model for heterochromatin formation, but the exact sequence of events for initiation remains to be elucidated. The essential factors involved include RNA transcribed from repeated sequences together with the methyltransferase Clr4. In addition, histone deacetylases, like Clr3, found in the SHREC complex are also necessary for transcriptional silencing. Clr2 is another crucial factor required for heterochromatin formation found in the SHREC complex. The function of Clr2 has been difficult to establish due to the lack of conserved domains or homology to proteins of known molecular function. Using a bioinformatics approach, three conserved motifs in Clr2 were identified, which contained amino acids important for transcriptional repression. Analysis of clr2 mutant strains revealed a major role for Clr2 in mating-type and rDNA silencing, and weaker effects on centromeric silencing. The effect on mating-type silencing showed variegation in several of the strains with mutated versions of Clr2 indicating an establishment or maintenance defect. Moreover, the critical amino acids in Clr2 were also necessary for transcriptional repression in a minimal system, by the tethering of Clr4 upstream of a reporter gene, inserted into the euchromatic part of the genome. Finally, in silico modeling suggested that the mutations in Clr2 cause disruption of secondary structures in the Clr2 protein. Identification of these critical amino acids in the protein provides a useful tool to explore the molecular mechanism behind the role of Clr2 in heterochromatin formation.
Conflict of interest statement
Figures

Similar articles
-
A novel type of silencing factor, Clr2, is necessary for transcriptional silencing at various chromosomal locations in the fission yeast Schizosaccharomyces pombe.Nucleic Acids Res. 2004 Aug 18;32(15):4421-8. doi: 10.1093/nar/gkh780. Print 2004. Nucleic Acids Res. 2004. PMID: 15317867 Free PMC article.
-
SHREC, an effector complex for heterochromatic transcriptional silencing.Cell. 2007 Feb 9;128(3):491-504. doi: 10.1016/j.cell.2006.12.035. Cell. 2007. PMID: 17289569
-
H3K9me-independent gene silencing in fission yeast heterochromatin by Clr5 and histone deacetylases.PLoS Genet. 2011 Jan 6;7(1):e1001268. doi: 10.1371/journal.pgen.1001268. PLoS Genet. 2011. PMID: 21253571 Free PMC article.
-
Studies on the mechanism of RNAi-dependent heterochromatin assembly.Cold Spring Harb Symp Quant Biol. 2006;71:461-71. doi: 10.1101/sqb.2006.71.044. Cold Spring Harb Symp Quant Biol. 2006. PMID: 17381328 Review.
-
RITS-connecting transcription, RNA interference, and heterochromatin assembly in fission yeast.Wiley Interdiscip Rev RNA. 2011 Sep-Oct;2(5):632-46. doi: 10.1002/wrna.80. Epub 2011 Mar 23. Wiley Interdiscip Rev RNA. 2011. PMID: 21823226 Free PMC article. Review.
Cited by
-
The binding of Chp2's chromodomain to methylated H3K9 is essential for Chp2's role in heterochromatin assembly in fission yeast.PLoS One. 2018 Aug 15;13(8):e0201101. doi: 10.1371/journal.pone.0201101. eCollection 2018. PLoS One. 2018. PMID: 30110338 Free PMC article.
-
Regulation of transcriptional silencing and chromodomain protein localization at centromeric heterochromatin by histone H3 tyrosine 41 phosphorylation in fission yeast.Nucleic Acids Res. 2018 Jan 9;46(1):189-202. doi: 10.1093/nar/gkx1010. Nucleic Acids Res. 2018. PMID: 29136238 Free PMC article.
-
Stress sensitivity of a fission yeast strain lacking histidine kinases is rescued by the ectopic expression of Chk1 from Candida albicans.Curr Genet. 2017 May;63(2):343-357. doi: 10.1007/s00294-016-0644-9. Epub 2016 Sep 9. Curr Genet. 2017. PMID: 27613427 Free PMC article.
-
SHREC Silences Heterochromatin via Distinct Remodeling and Deacetylation Modules.Mol Cell. 2016 Apr 21;62(2):207-221. doi: 10.1016/j.molcel.2016.03.016. Mol Cell. 2016. PMID: 27105116 Free PMC article.
-
Molecular fusion events in carcinogenic organisms: a bioinformatics study for the detection of fused proteins between viruses, bacteria and eukaryotes.EMBnet J. 2022 Apr;27:e1004. doi: 10.14806/ej.27.0.1004. Epub 2022 Apr 4. EMBnet J. 2022. PMID: 35464257 Free PMC article.
References
-
- Luger K, Mader AW, Richmond RK, Sargent DF, Richmond TJ (1997) Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature 389: 251–260. - PubMed
-
- Cam HP, Sugiyama T, Chen ES, Chen X, FitzGerald PC, et al. (2005) Comprehensive analysis of heterochromatin- and RNAi-mediated epigenetic control of the fission yeast genome. Nat Genet 37: 809–819. - PubMed
-
- Allshire RC, Nimmo ER, Ekwall K, Javerzat JP, Cranston G (1995) Mutations derepressing silent centromeric domains in fission yeast disrupt chromosome segregation. Genes Dev 9: 218–233. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
