

Gata3 drives development of RORγt+ group 3 innate lymphoid cells
- PMID: 24419270
- PMCID: PMC3920560
- DOI: 10.1084/jem.20131038
Gata3 drives development of RORγt+ group 3 innate lymphoid cells
Abstract
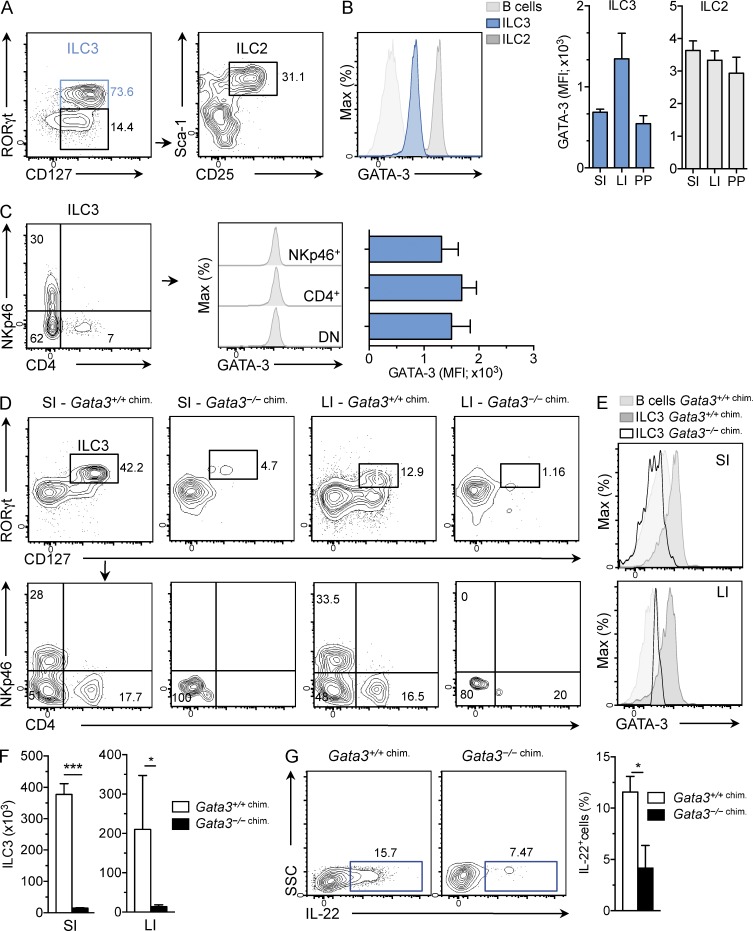
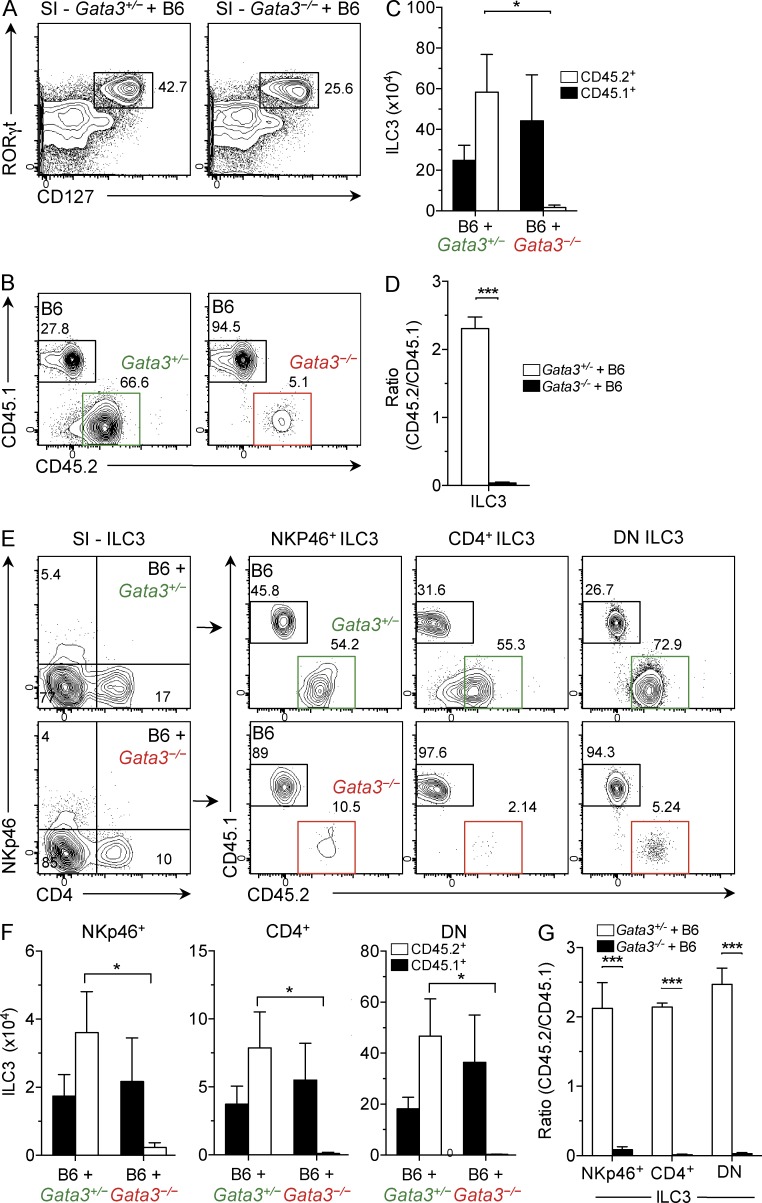
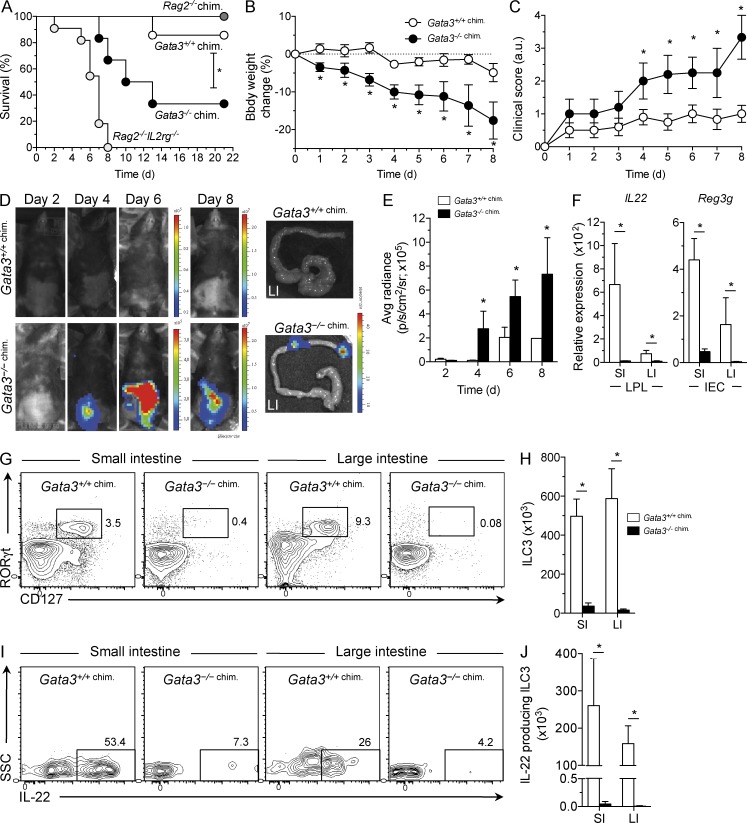
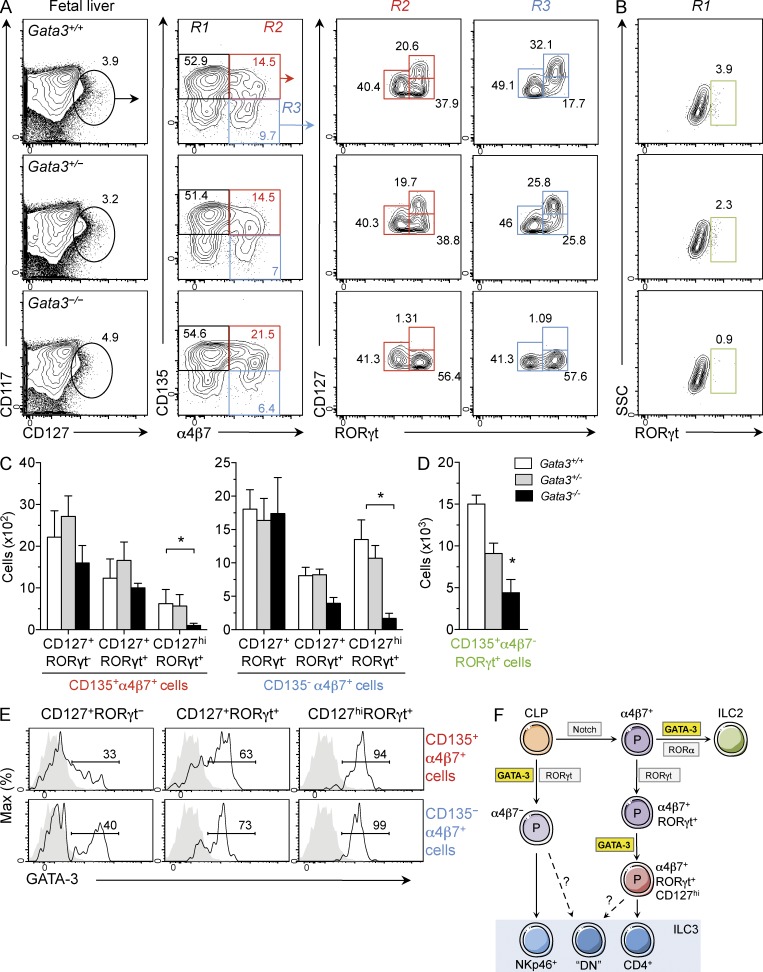
Group 3 innate lymphoid cells (ILC3) include IL-22-producing NKp46(+) cells and IL-17A/IL-22-producing CD4(+) lymphoid tissue inducerlike cells that express RORγt and are implicated in protective immunity at mucosal surfaces. Whereas the transcription factor Gata3 is essential for T cell and ILC2 development from hematopoietic stem cells (HSCs) and for IL-5 and IL-13 production by T cells and ILC2, the role for Gata3 in the generation or function of other ILC subsets is not known. We found that abundant GATA-3 protein is expressed in mucosa-associated ILC3 subsets with levels intermediate between mature B cells and ILC2. Chimeric mice generated with Gata3-deficient fetal liver hematopoietic precursors lack all intestinal RORγt(+) ILC3 subsets, and these mice show defective production of IL-22 early after infection with the intestinal pathogen Citrobacter rodentium, leading to impaired survival. Further analyses demonstrated that ILC3 development requires cell-intrinsic Gata3 expression in fetal liver hematopoietic precursors. Our results demonstrate that Gata3 plays a generalized role in ILC lineage determination and is critical for the development of gut RORγt(+) ILC3 subsets that maintain mucosal barrier homeostasis. These results further extend the paradigm of Gata3-dependent regulation of diversified innate ILC and adaptive T cell subsets.
Figures
Similar articles
-
TCF-1 controls ILC2 and NKp46+RORγt+ innate lymphocyte differentiation and protection in intestinal inflammation.J Immunol. 2013 Oct 15;191(8):4383-91. doi: 10.4049/jimmunol.1301228. Epub 2013 Sep 13. J Immunol. 2013. PMID: 24038093
-
Group 3 innate lymphoid cells continuously require the transcription factor GATA-3 after commitment.Nat Immunol. 2016 Feb;17(2):169-78. doi: 10.1038/ni.3318. Epub 2015 Nov 23. Nat Immunol. 2016. PMID: 26595886 Free PMC article.
-
Transcription Factor Bcl11b Controls Identity and Function of Mature Type 2 Innate Lymphoid Cells.Immunity. 2015 Aug 18;43(2):354-68. doi: 10.1016/j.immuni.2015.07.005. Epub 2015 Jul 28. Immunity. 2015. PMID: 26231117 Free PMC article.
-
[Roles of RORγt+ innate lymphoid cells in mucosal tissues of mouse and human].Nihon Rinsho Meneki Gakkai Kaishi. 2013;36(1):11-6. doi: 10.2177/jsci.36.11. Nihon Rinsho Meneki Gakkai Kaishi. 2013. PMID: 23445727 Review. Japanese.
-
Innate Lymphoid Cells and Intestinal Inflammatory Disorders.Int J Mol Sci. 2022 Feb 6;23(3):1856. doi: 10.3390/ijms23031856. Int J Mol Sci. 2022. PMID: 35163778 Free PMC article. Review.
Cited by
-
Innate lymphoid cell regulation of adaptive immunity.Immunology. 2016 Oct;149(2):123-30. doi: 10.1111/imm.12639. Epub 2016 Aug 16. Immunology. 2016. PMID: 27341319 Free PMC article. Review.
-
Identification of aceNKPs, a committed common progenitor population of the ILC1 and NK cell continuum.Proc Natl Acad Sci U S A. 2022 Dec 6;119(49):e2203454119. doi: 10.1073/pnas.2203454119. Epub 2022 Nov 29. Proc Natl Acad Sci U S A. 2022. PMID: 36442116 Free PMC article.
-
Identification of a novel lymphoid population in the murine epidermis.Sci Rep. 2015 Jul 30;5:12554. doi: 10.1038/srep12554. Sci Rep. 2015. PMID: 26223192 Free PMC article.
-
Retinoic Acid Differentially Regulates the Migration of Innate Lymphoid Cell Subsets to the Gut.Immunity. 2015 Jul 21;43(1):107-19. doi: 10.1016/j.immuni.2015.06.009. Epub 2015 Jun 30. Immunity. 2015. PMID: 26141583 Free PMC article.
-
The significance of long non-coding RNAs in the pathogenesis, diagnosis and treatment of inflammatory bowel disease.Precis Clin Med. 2023 Dec 11;6(4):pbad031. doi: 10.1093/pcmedi/pbad031. eCollection 2023 Dec. Precis Clin Med. 2023. PMID: 38163004 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
