

Neuronal interaural level difference response shifts are level-dependent in the rat auditory cortex
- PMID: 24335208
- PMCID: PMC3949225
- DOI: 10.1152/jn.00648.2013
Neuronal interaural level difference response shifts are level-dependent in the rat auditory cortex
Abstract
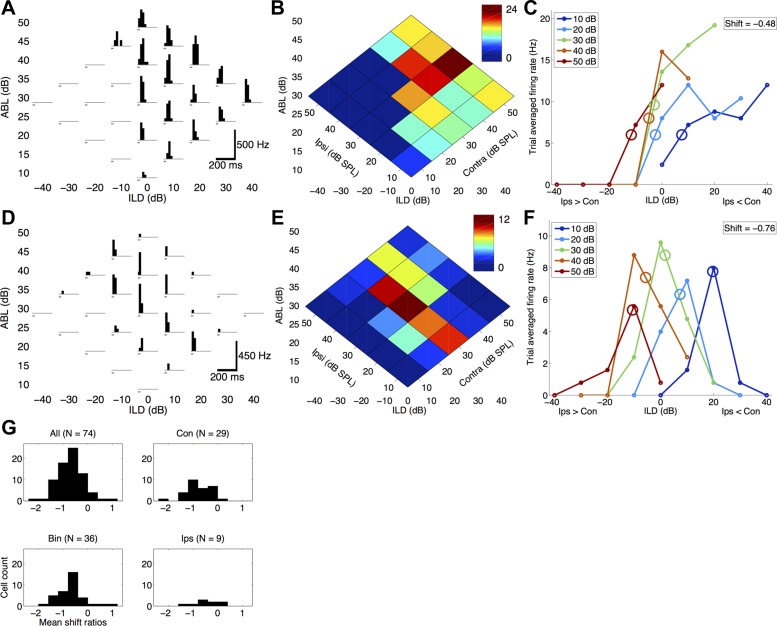
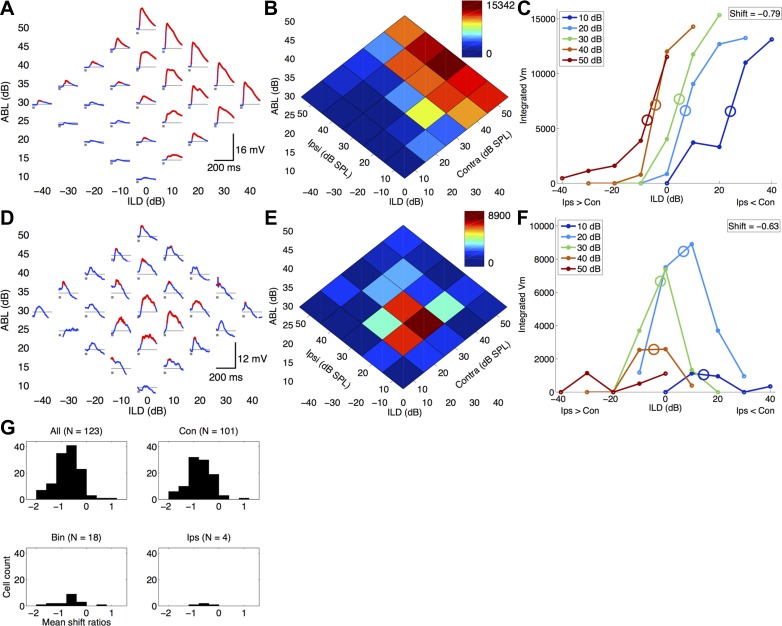
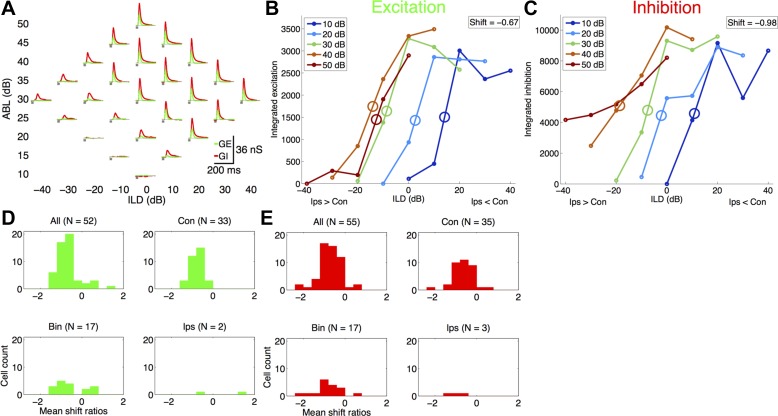
How does the brain accomplish sound localization with invariance to total sound level? Sensitivity to interaural level differences (ILDs) is first computed at the lateral superior olive (LSO) and is observed at multiple levels of the auditory pathway, including the central nucleus of inferior colliculus (ICC) and auditory cortex. In LSO, this ILD sensitivity is level-dependent, such that ILD response functions shift toward the ipsilateral (excitatory) ear with increasing sound level. Thus early in the processing pathway changes in firing rate could indicate changes in sound location, sound level, or both. In ICC, while ILD responses can shift toward either ear in individual neurons, there is no net ILD response shift at the population level. In behavioral studies of human sound localization acuity, ILD sensitivity is invariant to increasing sound levels. Level-invariant sound localization would suggest transformation in level sensitivity between LSO and perception of sound sources. Whether this transformation is completed at the level of the ICC or continued at higher levels remains unclear. It also remains unknown whether perceptual sound localization is level-invariant in rats, as it is in humans. We asked whether ILD sensitivity is level-invariant in rat auditory cortex. We performed single-unit and whole cell recordings in rat auditory cortex under ketamine anesthesia and measured responses to white noise bursts presented through sealed earphones at a range of ILDs. Surprisingly, we found that with increasing sound levels ILD responses shifted toward the ipsilateral ear (which is typically inhibitory), regardless of whether cells preferred ipsilateral, contralateral, or binaural stimuli. Voltage-clamp recordings suggest that synaptic inhibition does not contribute substantially to this transformation in level sensitivity. We conclude that the level invariance of ILD sensitivity seen in behavioral studies is not present in rat auditory cortex.
Keywords: auditory cortex; interaural level difference; level dependence; sound localization.
Figures
Similar articles
-
Synaptic mechanisms underlying interaural level difference selectivity in rat auditory cortex.J Neurophysiol. 2014 Nov 15;112(10):2561-71. doi: 10.1152/jn.00389.2014. Epub 2014 Sep 3. J Neurophysiol. 2014. PMID: 25185807 Free PMC article.
-
Slow Temporal Integration Enables Robust Neural Coding and Perception of a Cue to Sound Source Location.J Neurosci. 2016 Sep 21;36(38):9908-21. doi: 10.1523/JNEUROSCI.1421-16.2016. J Neurosci. 2016. PMID: 27656028 Free PMC article.
-
The coding of spatial location by single units in the lateral superior olive of the cat. I. Spatial receptive fields in azimuth.J Neurosci. 2002 Feb 15;22(4):1454-67. doi: 10.1523/JNEUROSCI.22-04-01454.2002. J Neurosci. 2002. PMID: 11850472 Free PMC article.
-
Sound localization.Handb Clin Neurol. 2015;129:99-116. doi: 10.1016/B978-0-444-62630-1.00006-8. Handb Clin Neurol. 2015. PMID: 25726265 Review.
-
The lateral superior olive: a functional role in sound source localization.Neuroscientist. 2003 Apr;9(2):127-43. doi: 10.1177/1073858403252228. Neuroscientist. 2003. PMID: 12708617 Review.
Cited by
-
Local and Global Spatial Organization of Interaural Level Difference and Frequency Preferences in Auditory Cortex.Cereb Cortex. 2018 Jan 1;28(1):350-369. doi: 10.1093/cercor/bhx295. Cereb Cortex. 2018. PMID: 29136122 Free PMC article.
-
Selectivity of Monaural Synaptic Inputs Underlying Binaural Auditory Information Integration in the Central Nucleus of Inferior Colliculus.Front Cell Neurosci. 2018 Oct 4;12:303. doi: 10.3389/fncel.2018.00303. eCollection 2018. Front Cell Neurosci. 2018. PMID: 30337856 Free PMC article.
-
Synaptic mechanisms underlying interaural level difference selectivity in rat auditory cortex.J Neurophysiol. 2014 Nov 15;112(10):2561-71. doi: 10.1152/jn.00389.2014. Epub 2014 Sep 3. J Neurophysiol. 2014. PMID: 25185807 Free PMC article.
-
State-dependent geometry of population activity in rat auditory cortex.Elife. 2019 Apr 10;8:e44526. doi: 10.7554/eLife.44526. Elife. 2019. PMID: 30969167 Free PMC article.
-
Opponent Coding of Sound Location (Azimuth) in Planum Temporale is Robust to Sound-Level Variations.Cereb Cortex. 2016 Jan;26(1):450-464. doi: 10.1093/cercor/bhv269. Epub 2015 Nov 5. Cereb Cortex. 2016. PMID: 26545618 Free PMC article.
References
-
- Altshuler MW, Comalli PE. Effect of stimulus intensity and frequency on median horizontal plane sound localization. J Aud Res 15: 262–265, 1975
-
- Barry PH. JPCalc, a software package for calculating liquid junction potential corrections in patch-clamp, intracellular, epithelial and bilayer measurements and for correcting junction potential measurements. J Neurosci Methods 51: 107–116, 1994 - PubMed
-
- Campbell RA, Doubell TP, Nodal FR, Schnupp JWH, King AJ. Interaural timing cues do not contribute to the map of space in the ferret superior colliculus: a virtual acoustic space study. J Neurophysiol 95: 242–254, 2006b - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources