

Death upon a kiss: mitochondrial outer membrane composition and organelle communication govern sensitivity to BAK/BAX-dependent apoptosis
- PMID: 24269152
- PMCID: PMC3947007
- DOI: 10.1016/j.chembiol.2013.10.009
Death upon a kiss: mitochondrial outer membrane composition and organelle communication govern sensitivity to BAK/BAX-dependent apoptosis
Abstract
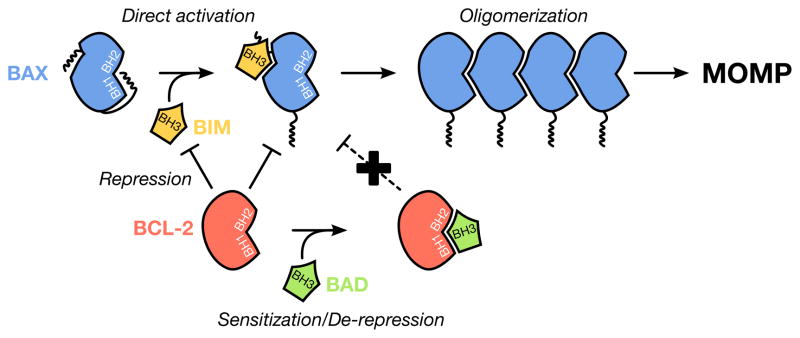
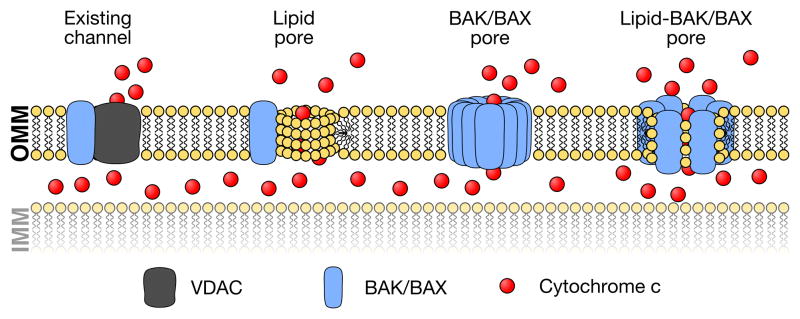
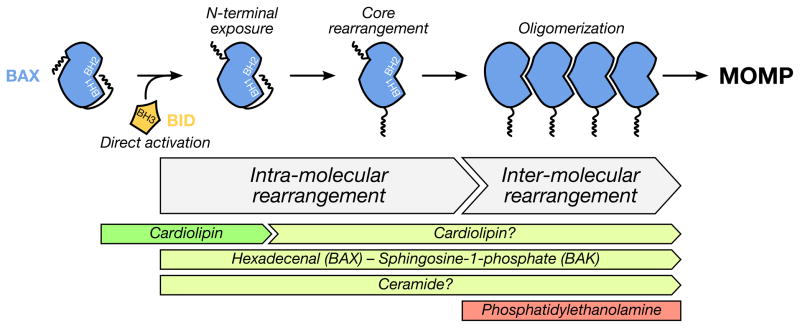
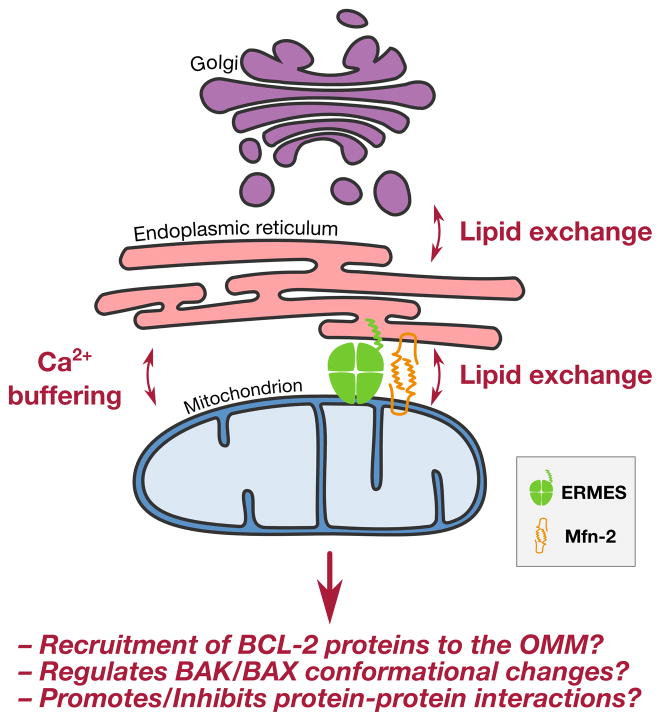
For stressed cells to induce the mitochondrial pathway of apoptosis, a cohort of pro-apoptotic BCL-2 proteins must collaborate with the outer mitochondrial membrane to permeabilize it. BAK and BAX are the two pro-apoptotic BCL-2 family members that are required for mitochondrial outer membrane permeabilization. While biochemical and structural insights of BAK/BAX function have expanded in recent years, very little is known about the role of the outer mitochondrial membrane in regulating BAK/BAX activity. In this review, we will highlight the impact of mitochondrial composition (both protein and lipid) and mitochondrial interactions with cellular organelles on BAK/BAX function and cellular commitment to apoptosis. A better understanding of how BAK/BAX and mitochondrial biology are mechanistically linked will likely reveal novel insights into homeostatic and pathological mechanisms associated with apoptosis.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures
Similar articles
-
Bax, Bak and beyond - mitochondrial performance in apoptosis.FEBS J. 2018 Feb;285(3):416-431. doi: 10.1111/febs.14186. Epub 2017 Sep 4. FEBS J. 2018. PMID: 28755482 Review.
-
Building blocks of the apoptotic pore: how Bax and Bak are activated and oligomerize during apoptosis.Cell Death Differ. 2014 Feb;21(2):196-205. doi: 10.1038/cdd.2013.139. Epub 2013 Oct 25. Cell Death Differ. 2014. PMID: 24162660 Free PMC article. Review.
-
Physiological and Pharmacological Control of BAK, BAX, and Beyond.Trends Cell Biol. 2016 Dec;26(12):906-917. doi: 10.1016/j.tcb.2016.07.002. Epub 2016 Aug 4. Trends Cell Biol. 2016. PMID: 27498846 Free PMC article. Review.
-
Apoptotic pore formation is associated with in-plane insertion of Bak or Bax central helices into the mitochondrial outer membrane.Proc Natl Acad Sci U S A. 2014 Sep 30;111(39):E4076-85. doi: 10.1073/pnas.1415142111. Epub 2014 Sep 16. Proc Natl Acad Sci U S A. 2014. PMID: 25228770 Free PMC article.
-
Structure-based modeling of turnover of Bcl-2 family proteins bound to voltage-dependent anion channel 2 (VDAC2): Implications for the mechanisms of proapoptotic activation of Bak and Bax in vivo.Comput Biol Chem. 2020 Apr;85:107203. doi: 10.1016/j.compbiolchem.2020.107203. Epub 2020 Jan 13. Comput Biol Chem. 2020. PMID: 31981967
Cited by
-
Conformational Heterogeneity of Bax Helix 9 Dimer for Apoptotic Pore Formation.Sci Rep. 2016 Jul 6;6:29502. doi: 10.1038/srep29502. Sci Rep. 2016. PMID: 27381287 Free PMC article.
-
Putting the pieces together: How is the mitochondrial pathway of apoptosis regulated in cancer and chemotherapy?Cancer Metab. 2014 Oct 6;2:16. doi: 10.1186/2049-3002-2-16. eCollection 2014. Cancer Metab. 2014. PMID: 25621172 Free PMC article. Review.
-
Chorein addiction in VPS13A overexpressing rhabdomyosarcoma cells.Oncotarget. 2015 Apr 30;6(12):10309-19. doi: 10.18632/oncotarget.3582. Oncotarget. 2015. PMID: 25871399 Free PMC article.
-
Indomethacin impairs mitochondrial dynamics by activating the PKCζ-p38-DRP1 pathway and inducing apoptosis in gastric cancer and normal mucosal cells.J Biol Chem. 2019 May 17;294(20):8238-8258. doi: 10.1074/jbc.RA118.004415. Epub 2019 Apr 2. J Biol Chem. 2019. PMID: 30940726 Free PMC article.
-
Molecular effects of genistein, as a potential anticancer agent, on CXCR-4 and VEGF pathway in acute lymphoblastic leukemia.Mol Biol Rep. 2022 Jun;49(6):4161-4170. doi: 10.1007/s11033-022-07163-0. Epub 2022 May 24. Mol Biol Rep. 2022. PMID: 35608747
References
-
- Antonsson B, Montessuit S, Sanchez B, Martinou JC. Bax is present as a high molecular weight oligomer/complex in the mitochondrial membrane of apoptotic cells. J Biol Chem. 2001;276:11615–11623. - PubMed
-
- Basañez G, Sharpe JC, Galanis J, Brandt TB, Hardwick JM, Zimmerberg J. Bax-type apoptotic proteins porate pure lipid bilayers through a mechanism sensitive to intrinsic monolayer curvature. J Biol Chem. 2002;277:49360–49365. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
