

Cyclin-dependent kinase complexes in developing maize endosperm: evidence for differential expression and functional specialization
- PMID: 24240479
- PMCID: PMC3902077
- DOI: 10.1007/s00425-013-1990-1
Cyclin-dependent kinase complexes in developing maize endosperm: evidence for differential expression and functional specialization
Abstract
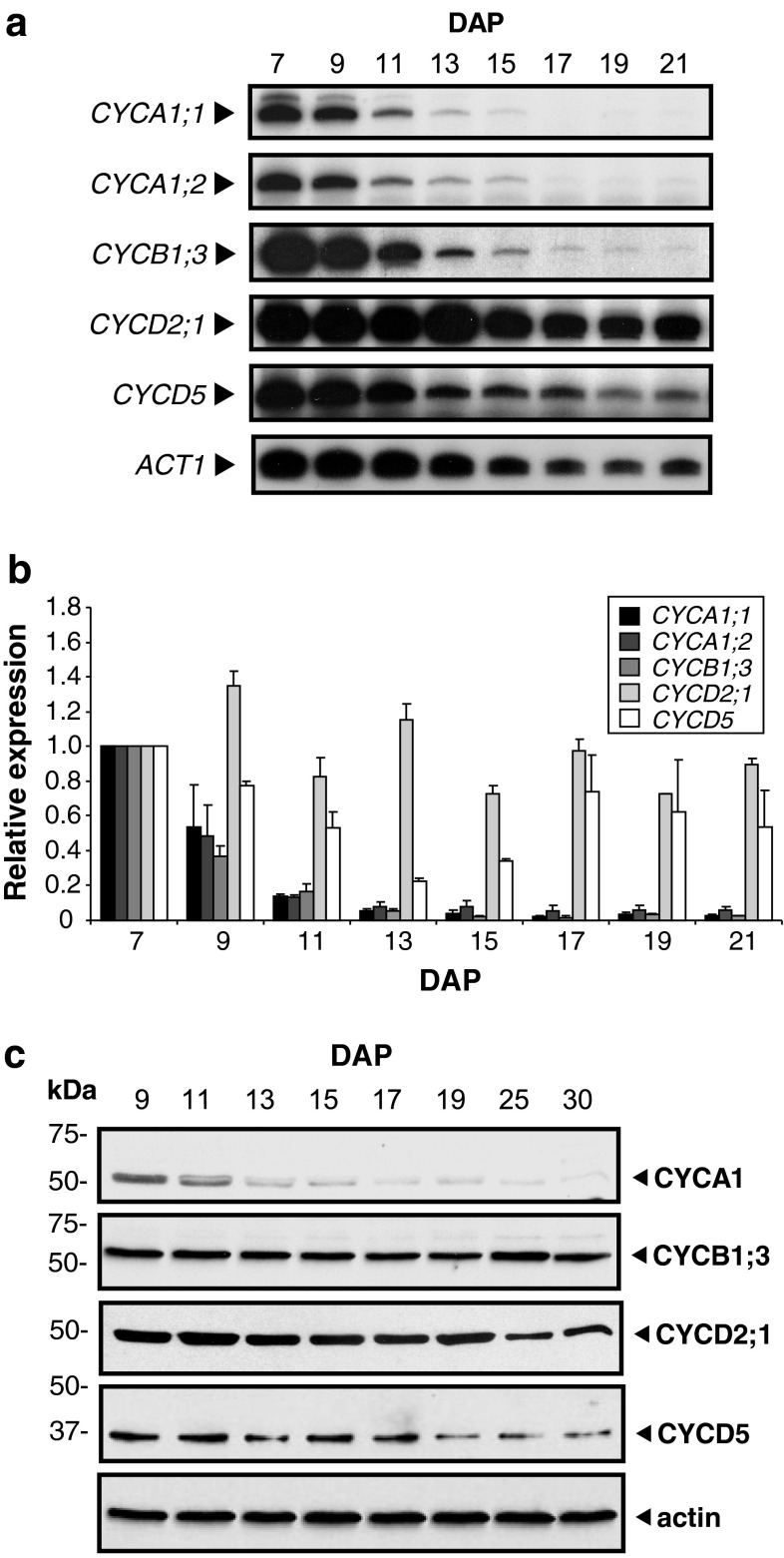
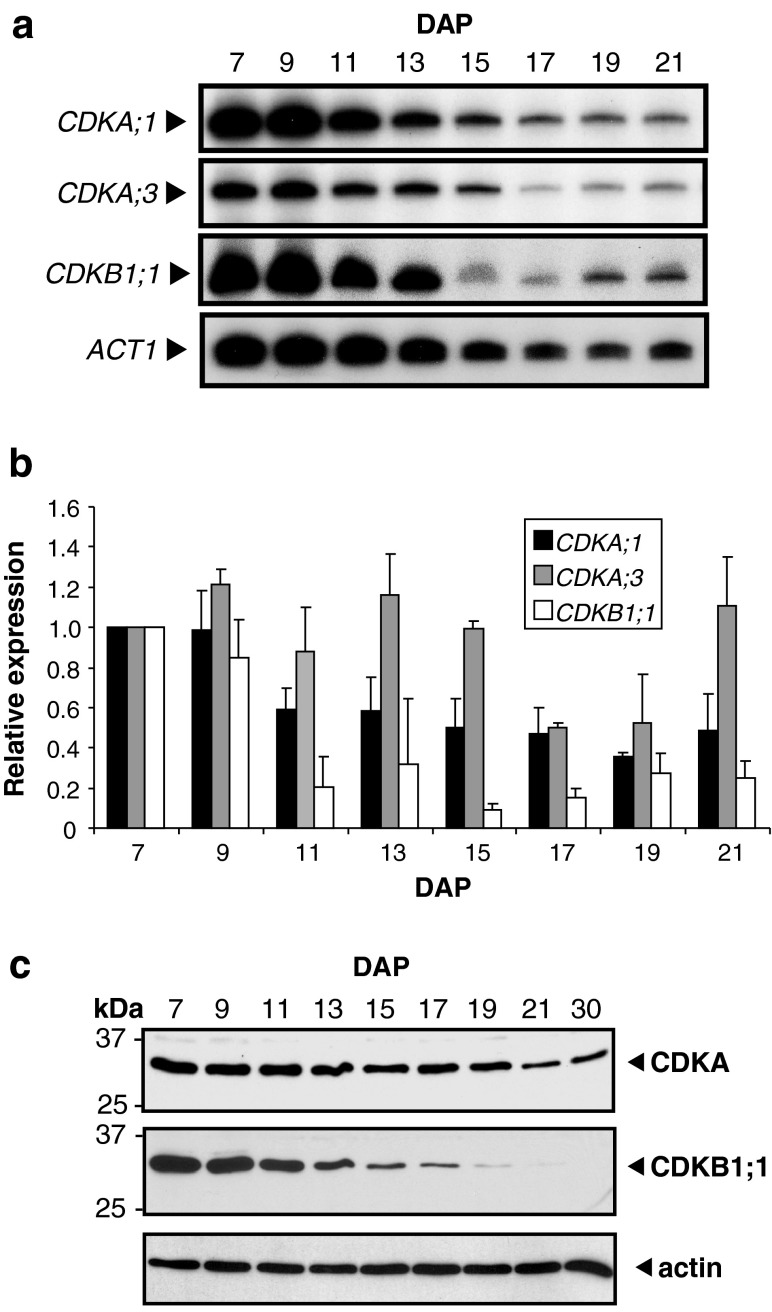
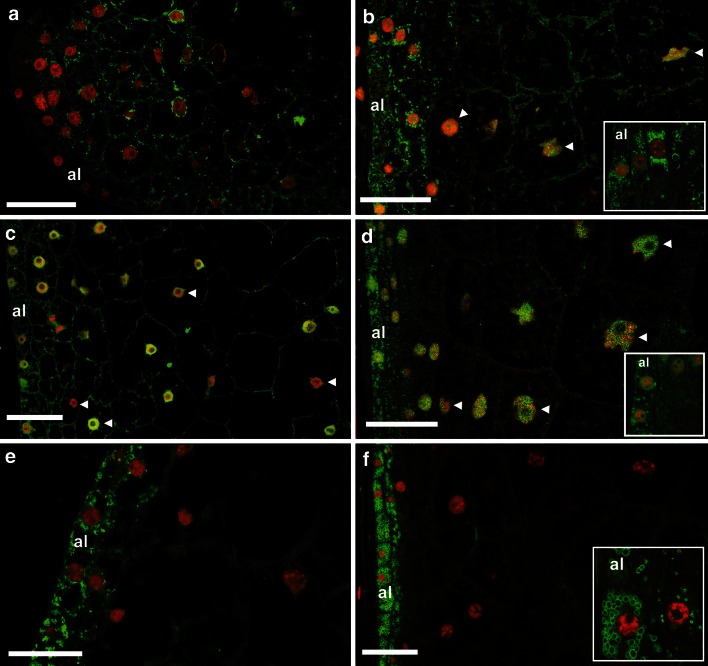
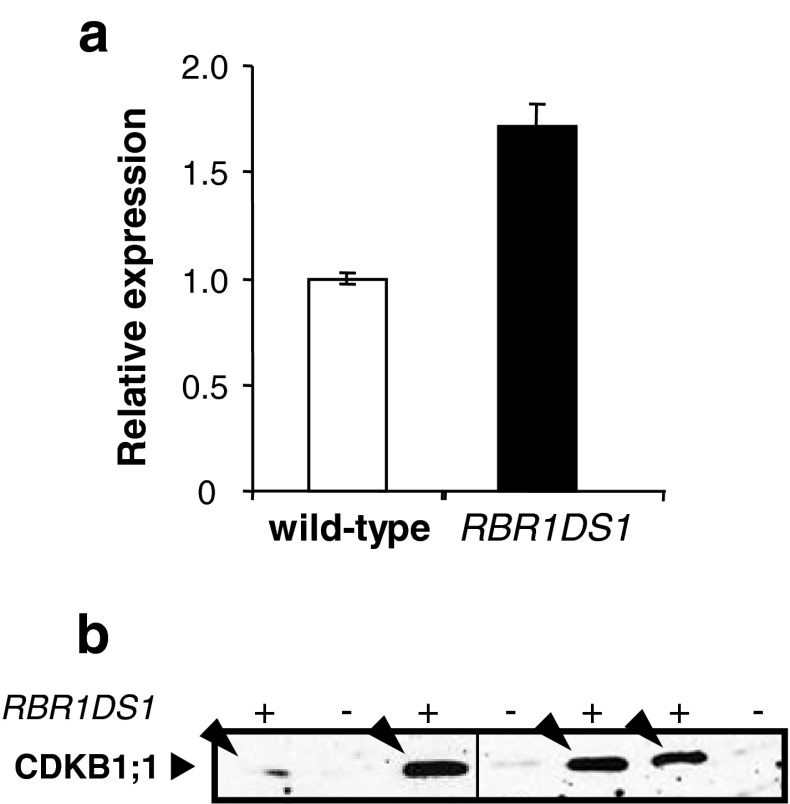
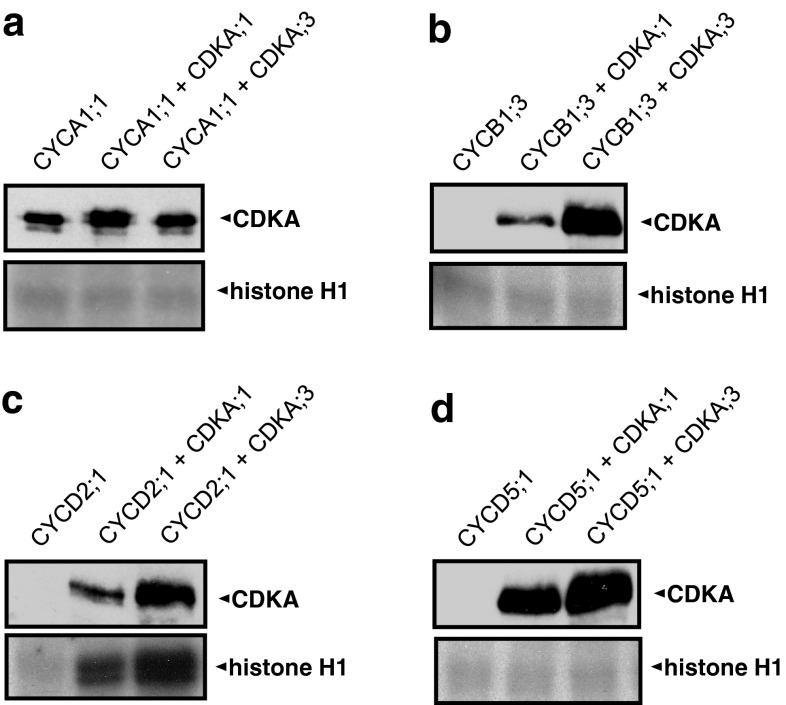
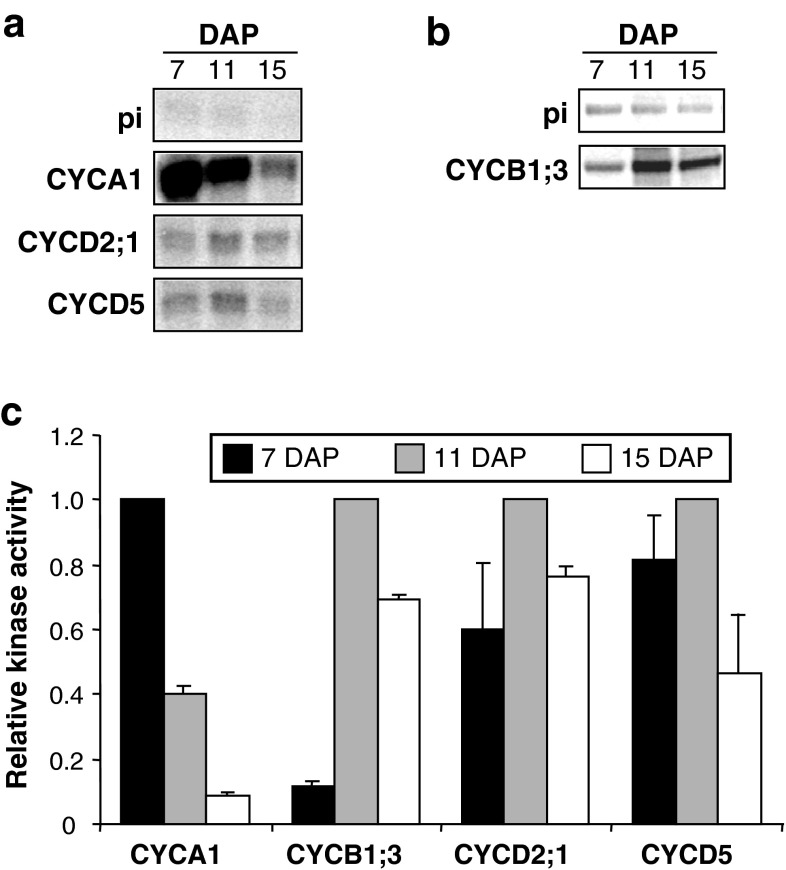
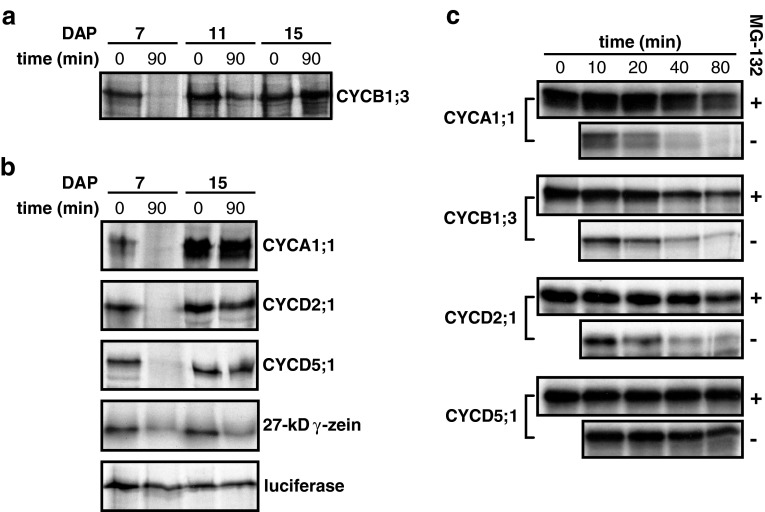
Endosperm development in maize (Zea mays L.) and related cereals comprises a cell proliferation stage followed by a period of rapid growth coupled to endoreduplication. Regulation of the cell cycle in developing endosperm is poorly understood. We have characterized various subunits of cyclin-dependent kinase (CDK) complexes, master cell cycle regulators in all eukaryotes. A-, B-, and D-type cyclins as well as A- and B-type cyclin-dependent kinases were characterized with respect to their RNA and protein expression profiles. Two main patterns were identified: one showing expression throughout endosperm development, and another characterized by a sharp down-regulation with the onset of endoreduplication. Cyclin CYCB1;3 and CYCD2;1 proteins were distributed in the cytoplasm and nucleus of cells throughout the endosperm, while cyclin CYCD5 protein was localized in the cytoplasm of peripheral cells. CDKB1;1 expression was strongly associated with cell proliferation. Expression and cyclin-binding patterns suggested that CDKA;1 and CDKA;3 are at least partially redundant. The kinase activity associated with the cyclin CYCA1 was highest during the mitotic stage of development, while that associated with CYCB1;3, CYCD2;1 and CYCD5 peaked at the mitosis-to-endoreduplication transition. A-, B- and D-type cyclins were more resistant to proteasome-dependent degradation in endoreduplicating than in mitotic endosperm extracts. These results indicated that endosperm development is characterized by differential expression and activity of specific cyclins and CDKs, and suggested that endoreduplication is associated with reduced cyclin proteolysis via the ubiquitin-proteasome pathway.
Figures
Similar articles
-
Expression, regulation and activity of a B2-type cyclin in mitotic and endoreduplicating maize endosperm.Front Plant Sci. 2014 Oct 17;5:561. doi: 10.3389/fpls.2014.00561. eCollection 2014. Front Plant Sci. 2014. PMID: 25368625 Free PMC article.
-
A dominant negative mutant of cyclin-dependent kinase A reduces endoreduplication but not cell size or gene expression in maize endosperm.Plant Cell. 2004 Jul;16(7):1854-69. doi: 10.1105/tpc.022178. Epub 2004 Jun 18. Plant Cell. 2004. PMID: 15208390 Free PMC article.
-
The cyclin-dependent kinase inhibitor KRP2 controls the onset of the endoreduplication cycle during Arabidopsis leaf development through inhibition of mitotic CDKA;1 kinase complexes.Plant Cell. 2005 Jun;17(6):1723-36. doi: 10.1105/tpc.105.032383. Epub 2005 Apr 29. Plant Cell. 2005. PMID: 15863515 Free PMC article.
-
Investigating the hows and whys of DNA endoreduplication.J Exp Bot. 2001 Feb;52(355):183-92. J Exp Bot. 2001. PMID: 11283162 Review.
-
The contribution of cell cycle regulation to endosperm development.Sex Plant Reprod. 2009 Dec;22(4):207-19. doi: 10.1007/s00497-009-0105-4. Epub 2009 Jul 26. Sex Plant Reprod. 2009. PMID: 20033442 Review.
Cited by
-
Characterization key genes of Arabidopsis seedlings in response to β-caryophyllene, eugenol using combined transcriptome and WGCN analysis.Front Plant Sci. 2024 Jan 4;14:1295779. doi: 10.3389/fpls.2023.1295779. eCollection 2023. Front Plant Sci. 2024. PMID: 38239209 Free PMC article.
-
Dissection of Developmental Programs and Regulatory Modules Directing Endosperm Transfer Cell and Aleurone Identity in the Syncytial Endosperm of Barley.Plants (Basel). 2023 Apr 10;12(8):1594. doi: 10.3390/plants12081594. Plants (Basel). 2023. PMID: 37111818 Free PMC article.
-
Combating Dual Challenges in Maize Under High Planting Density: Stem Lodging and Kernel Abortion.Front Plant Sci. 2021 Nov 2;12:699085. doi: 10.3389/fpls.2021.699085. eCollection 2021. Front Plant Sci. 2021. PMID: 34868101 Free PMC article. Review.
-
Cell cycle control and seed development.Front Plant Sci. 2014 Sep 23;5:493. doi: 10.3389/fpls.2014.00493. eCollection 2014. Front Plant Sci. 2014. PMID: 25295050 Free PMC article. Review.
-
Transcriptome Dynamics during Maize Endosperm Development.PLoS One. 2016 Oct 3;11(10):e0163814. doi: 10.1371/journal.pone.0163814. eCollection 2016. PLoS One. 2016. PMID: 27695101 Free PMC article.
References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Boudolf V, Vlieghe K, Beemster GTS, Magyar Z, Acosta JAT, Maes S, Van Der Schueren E, Inzé D, De Veylder L. The plant-specific cyclin-dependent kinase CDKB1;1 and transcription factor E2Fa-DPa control the balance of mitotically dividing and endoreduplicating cells in Arabidopsis. Plant Cell. 2004;16:2683–2692. doi: 10.1105/tpc.104.024398. - DOI - PMC - PubMed
-
- Boudolf V, Lammen T, Boruc J, Van Leene J, Van Den Daele H, Maes S, Van Isterdael G, Russinova E, Kondorosi E, Witters E, De Jaeger G, Inzé D, De Veylder L. CDKB1;1 forms a functional complex with CYCA2;3 to suppress endocycle onset. Plant Physiol. 2009;150:1482–1493. doi: 10.1104/pp.109.140269. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
