

Mechanism of HIV-1 neutralization by antibodies targeting a membrane-proximal region of gp41
- PMID: 24227838
- PMCID: PMC3911647
- DOI: 10.1128/JVI.02664-13
Mechanism of HIV-1 neutralization by antibodies targeting a membrane-proximal region of gp41
Abstract
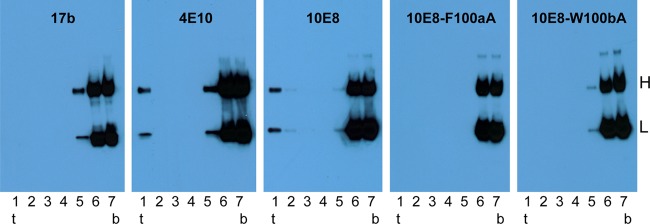
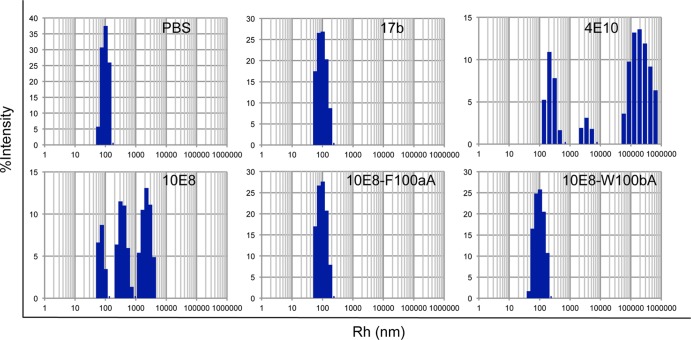
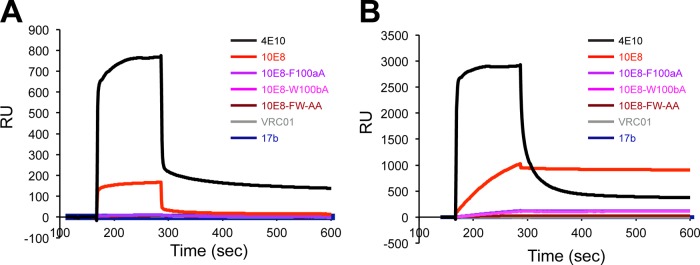
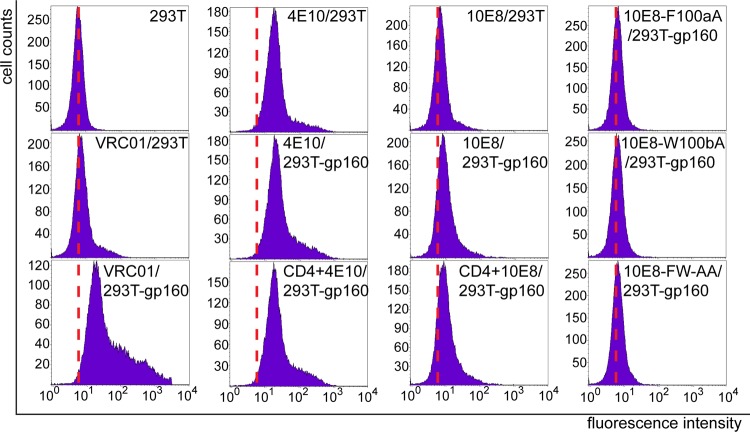
Induction of broadly neutralizing antibodies (bNAbs) is an important goal for HIV-1 vaccine development. Two autoreactive bNAbs, 2F5 and 4E10, recognize a conserved region on the HIV-1 envelope glycoprotein gp41 adjacent to the viral membrane known as the membrane-proximal external region (MPER). They block viral infection by targeting a fusion-intermediate conformation of gp41, assisted by an additional interaction with the viral membrane. Another MPER-specific antibody, 10E8, has recently been reported to neutralize HIV-1 with potency and breadth much greater than those of 2F5 or 4E10, but it appeared not to bind phospholipids and might target the untriggered envelope spikes, raising the hope that the MPER could be harnessed for vaccine design without major immunological concerns. Here, we show by three independent approaches that 10E8 indeed binds lipid bilayers through two hydrophobic residues in its CDR H3 (third heavy-chain complementarity-determining region). Its weak affinity for membranes in general and preference for cholesterol-rich membranes may account for its great neutralization potency, as it is less likely than other MPER-specific antibodies to bind cellular membranes nonspecifically. 10E8 binds with high affinity to a construct mimicking the fusion intermediate of gp41 but fails to recognize the envelope trimers representing the untriggered conformation. Moreover, we can improve the potency of 4E10 without affecting its binding to gp41 by a modification of its lipid-interacting CDR H3. These results reveal a general mechanism of HIV-1 neutralization by MPER-specific antibodies that involves interactions with viral lipids.
Figures

Similar articles
-
Interaction of anti-HIV type 1 antibody 2F5 with phospholipid bilayers and its relevance for the mechanism of virus neutralization.AIDS Res Hum Retroviruses. 2011 Aug;27(8):863-76. doi: 10.1089/AID.2010.0265. Epub 2011 Jan 15. AIDS Res Hum Retroviruses. 2011. PMID: 21142698
-
Developmental Pathway of the MPER-Directed HIV-1-Neutralizing Antibody 10E8.PLoS One. 2016 Jun 14;11(6):e0157409. doi: 10.1371/journal.pone.0157409. eCollection 2016. PLoS One. 2016. PMID: 27299673 Free PMC article.
-
Structural basis for HIV-1 neutralization by 2F5-like antibodies m66 and m66.6.J Virol. 2014 Mar;88(5):2426-41. doi: 10.1128/JVI.02837-13. Epub 2013 Dec 11. J Virol. 2014. PMID: 24335316 Free PMC article.
-
Antigp41 membrane proximal external region antibodies and the art of using the membrane for neutralization.Curr Opin HIV AIDS. 2017 May;12(3):250-256. doi: 10.1097/COH.0000000000000364. Curr Opin HIV AIDS. 2017. PMID: 28422789 Review.
-
Neutralizing Antibodies Targeting HIV-1 gp41.Viruses. 2020 Oct 23;12(11):1210. doi: 10.3390/v12111210. Viruses. 2020. PMID: 33114242 Free PMC article. Review.
Cited by
-
Inhibitory Effect of Individual or Combinations of Broadly Neutralizing Antibodies and Antiviral Reagents against Cell-Free and Cell-to-Cell HIV-1 Transmission.J Virol. 2015 Aug;89(15):7813-28. doi: 10.1128/JVI.00783-15. Epub 2015 May 20. J Virol. 2015. PMID: 25995259 Free PMC article.
-
Molecular recognition of the native HIV-1 MPER revealed by STED microscopy of single virions.Nat Commun. 2019 Jan 8;10(1):78. doi: 10.1038/s41467-018-07962-9. Nat Commun. 2019. PMID: 30622256 Free PMC article.
-
Structure and Recognition of a Novel HIV-1 gp120-gp41 Interface Antibody that Caused MPER Exposure through Viral Escape.PLoS Pathog. 2017 Jan 11;13(1):e1006074. doi: 10.1371/journal.ppat.1006074. eCollection 2017 Jan. PLoS Pathog. 2017. PMID: 28076415 Free PMC article.
-
A heterologous prime-boosting strategy with replicating Vaccinia virus vectors and plant-produced HIV-1 Gag/dgp41 virus-like particles.Virology. 2017 Jul;507:242-256. doi: 10.1016/j.virol.2017.04.008. Epub 2017 Apr 28. Virology. 2017. PMID: 28458036 Free PMC article.
-
Optimization of the Solubility of HIV-1-Neutralizing Antibody 10E8 through Somatic Variation and Structure-Based Design.J Virol. 2016 Jun 10;90(13):5899-5914. doi: 10.1128/JVI.03246-15. Print 2016 Jul 1. J Virol. 2016. PMID: 27053554 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
