

Reselection of a genomic upstream open reading frame in mouse hepatitis coronavirus 5'-untranslated-region mutants
- PMID: 24173235
- PMCID: PMC3911626
- DOI: 10.1128/JVI.02831-13
Reselection of a genomic upstream open reading frame in mouse hepatitis coronavirus 5'-untranslated-region mutants
Abstract
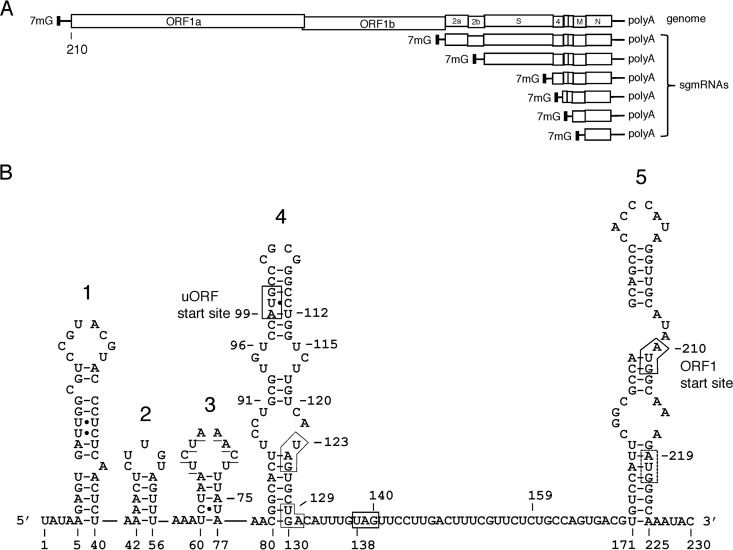
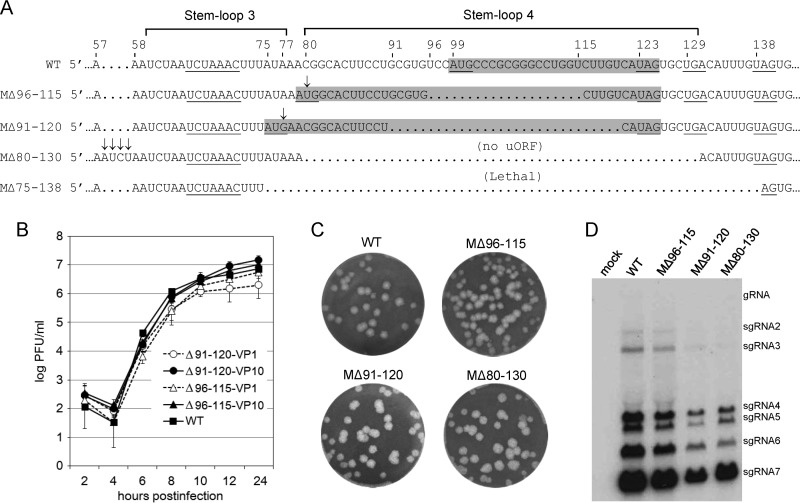
An AUG-initiated upstream open reading frame (uORF) encoding a potential polypeptide of 3 to 13 amino acids (aa) is found within the 5' untranslated region (UTR) of >75% of coronavirus genomes based on 38 reference strains. Potential CUG-initiated uORFs are also found in many strains. The AUG-initiated uORF is presumably translated following genomic 5'-end cap-dependent ribosomal scanning, but its function is unknown. Here, in a reverse-genetics study with mouse hepatitis coronavirus, the following were observed. (i) When the uORF AUG-initiating codon was replaced with a UAG stop codon along with a U112A mutation to maintain a uORF-harboring stem-loop 4 structure, an unimpaired virus with wild-type (WT) growth kinetics was recovered. However, reversion was found at all mutated sites within five virus passages. (ii) When the uORF was fused with genomic (main) ORF1 by converting three in-frame stop codons to nonstop codons, a uORF-ORF1 fusion protein was made, and virus replicated at WT levels. However, a frameshifting G insertion at virus passage 7 established a slightly 5'-extended original uORF. (iii) When uAUG-eliminating deletions of 20, 30, or 51 nucleotides (nt) were made within stem-loop 4, viable but debilitated virus was recovered. However, a C80U mutation in the first mutant and an A77G mutation in the second appeared by passage 10, which generated alternate uORFs that correlated with restored WT growth kinetics. In vitro, the uORF-disrupting nondeletion mutants showed enhanced translation of the downstream ORF1 compared with the WT. These results together suggest that the uORF represses ORF1 translation yet plays a beneficial but nonessential role in coronavirus replication in cell culture.
Figures
Similar articles
-
An optimal cis-replication stem-loop IV in the 5' untranslated region of the mouse coronavirus genome extends 16 nucleotides into open reading frame 1.J Virol. 2011 Jun;85(11):5593-605. doi: 10.1128/JVI.00263-11. Epub 2011 Mar 23. J Virol. 2011. PMID: 21430057 Free PMC article.
-
Negative and translation termination-dependent positive control of FLI-1 protein synthesis by conserved overlapping 5' upstream open reading frames in Fli-1 mRNA.Mol Cell Biol. 2000 May;20(9):2959-69. doi: 10.1128/MCB.20.9.2959-2969.2000. Mol Cell Biol. 2000. PMID: 10757781 Free PMC article.
-
Temperature-dependent regulation of upstream open reading frame translation in S. cerevisiae.BMC Biol. 2019 Dec 6;17(1):101. doi: 10.1186/s12915-019-0718-5. BMC Biol. 2019. PMID: 31810458 Free PMC article.
-
Translational Regulation by Upstream Open Reading Frames and Human Diseases.Adv Exp Med Biol. 2019;1157:99-116. doi: 10.1007/978-3-030-19966-1_5. Adv Exp Med Biol. 2019. PMID: 31342439 Review.
-
Specific Increase of Protein Levels by Enhancing Translation Using Antisense Oligonucleotides Targeting Upstream Open Frames.Adv Exp Med Biol. 2017;983:129-146. doi: 10.1007/978-981-10-4310-9_9. Adv Exp Med Biol. 2017. PMID: 28639196 Review.
Cited by
-
The structure and functions of coronavirus genomic 3' and 5' ends.Virus Res. 2015 Aug 3;206:120-33. doi: 10.1016/j.virusres.2015.02.025. Epub 2015 Feb 28. Virus Res. 2015. PMID: 25736566 Free PMC article. Review.
-
The interface between coronaviruses and host cell RNA biology: Novel potential insights for future therapeutic intervention.Wiley Interdiscip Rev RNA. 2020 Sep;11(5):e1614. doi: 10.1002/wrna.1614. Epub 2020 Jul 7. Wiley Interdiscip Rev RNA. 2020. PMID: 32638509 Free PMC article. Review.
-
Viral and Cellular mRNA Translation in Coronavirus-Infected Cells.Adv Virus Res. 2016;96:165-192. doi: 10.1016/bs.aivir.2016.08.001. Epub 2016 Sep 10. Adv Virus Res. 2016. PMID: 27712623 Free PMC article. Review.
-
Lessons Learned and Yet-to-Be Learned on the Importance of RNA Structure in SARS-CoV-2 Replication.Microbiol Mol Biol Rev. 2022 Sep 21;86(3):e0005721. doi: 10.1128/mmbr.00057-21. Epub 2022 Jul 7. Microbiol Mol Biol Rev. 2022. PMID: 35862724 Free PMC article. Review.
-
Host-like RNA Elements Regulate Virus Translation.Viruses. 2024 Mar 20;16(3):468. doi: 10.3390/v16030468. Viruses. 2024. PMID: 38543832 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
