

'Clustering' SIRPα into the plasma membrane lipid microdomains is required for activated monocytes and macrophages to mediate effective cell surface interactions with CD47
- PMID: 24143245
- PMCID: PMC3797048
- DOI: 10.1371/journal.pone.0077615
'Clustering' SIRPα into the plasma membrane lipid microdomains is required for activated monocytes and macrophages to mediate effective cell surface interactions with CD47
Abstract
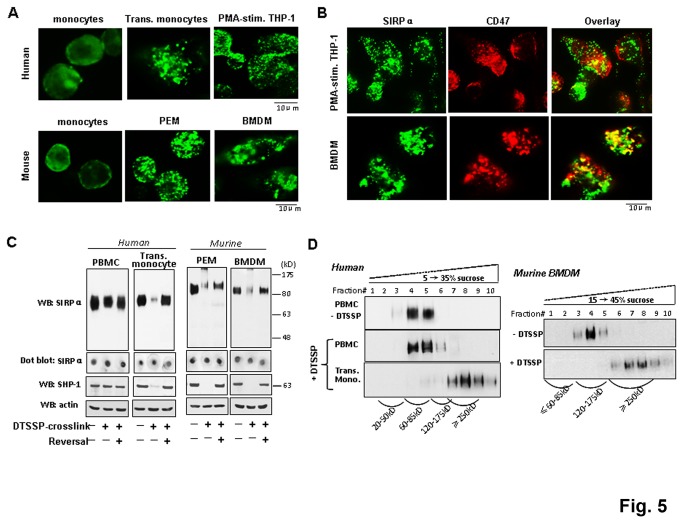
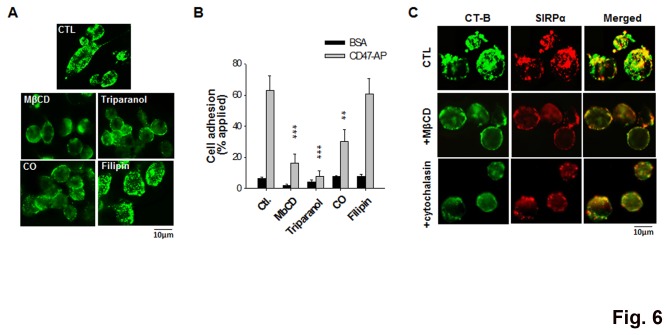
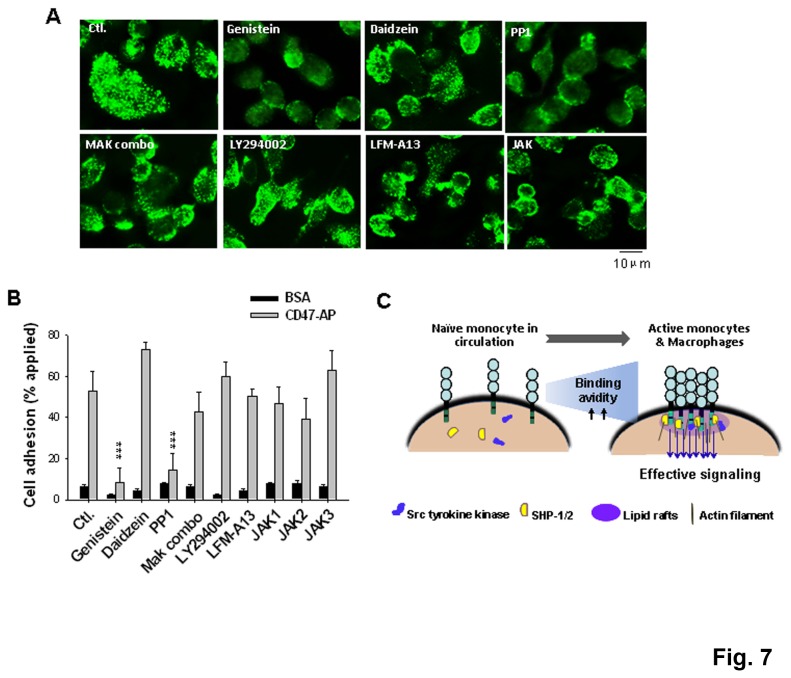
SIRPα, an ITIMs-containing signaling receptor, negatively regulates leukocyte responses through extracellular interactions with CD47. However, the dynamics of SIRPα-CD47 interactions on the cell surface and the governing mechanisms remain unclear. Here we report that while the purified SIRPα binds to CD47 and that SIRPα is expressed on monocytes and monocytic THP-1 or U937, these SIRPα are ineffective to mediate cell binding to immobilized CD47. However, cell binding to CD47 is significantly enhanced when monocytes transmigrating across endothelia, or being differentiated into macrophages. Cell surface labeling reveals SIRPα to be diffused on naïve monocytes but highly clustered on transmigrated monocytes and macrophages. Protein crosslink and equilibrium centrifugation confirm that SIRPα in the latter cells forms oligomerized complexes resulting in increased avidity for CD47 binding. Furthermore, formation of SIRPα complexes/clusters requires the plasma membrane 'lipid rafts' and the activity of Src family kinase during macrophage differentiation. These results together suggest that 'clustering' SIRPα into plasma membrane microdomains is essential for activated monocytes and macrophages to effectively interact with CD47 and initiate intracellular signaling.
Conflict of interest statement
Figures
Similar articles
-
Loss of Cell Surface CD47 Clustering Formation and Binding Avidity to SIRPα Facilitate Apoptotic Cell Clearance by Macrophages.J Immunol. 2015 Jul 15;195(2):661-71. doi: 10.4049/jimmunol.1401719. Epub 2015 Jun 17. J Immunol. 2015. PMID: 26085683 Free PMC article.
-
Exosome-SIRPα, a CD47 blockade increases cancer cell phagocytosis.Biomaterials. 2017 Mar;121:121-129. doi: 10.1016/j.biomaterials.2017.01.004. Epub 2017 Jan 3. Biomaterials. 2017. PMID: 28086180
-
Functional CD47/signal regulatory protein alpha (SIRP(alpha)) interaction is required for optimal human T- and natural killer- (NK) cell homeostasis in vivo.Proc Natl Acad Sci U S A. 2011 Aug 9;108(32):13224-9. doi: 10.1073/pnas.1101398108. Epub 2011 Jul 25. Proc Natl Acad Sci U S A. 2011. PMID: 21788504 Free PMC article.
-
The regulation of CD47-SIRPα signaling axis by microRNAs in combination with conventional cytotoxic drugs together with the help of nano-delivery: a choice for therapy?Mol Biol Rep. 2021 Jul;48(7):5707-5722. doi: 10.1007/s11033-021-06547-y. Epub 2021 Jul 17. Mol Biol Rep. 2021. PMID: 34275112 Review.
-
The CD47-SIRPα signaling axis as an innate immune checkpoint in cancer.Immunol Rev. 2017 Mar;276(1):145-164. doi: 10.1111/imr.12527. Immunol Rev. 2017. PMID: 28258703 Review.
Cited by
-
Cd47-Sirpα interaction and IL-10 constrain inflammation-induced macrophage phagocytosis of healthy self-cells.Proc Natl Acad Sci U S A. 2016 Sep 13;113(37):E5434-43. doi: 10.1073/pnas.1521069113. Epub 2016 Aug 30. Proc Natl Acad Sci U S A. 2016. PMID: 27578867 Free PMC article.
-
Using the Jurkat reporter T cell line for evaluating the functionality of novel chimeric antigen receptors.Front Mol Med. 2023 Feb 22;3:1070384. doi: 10.3389/fmmed.2023.1070384. eCollection 2023. Front Mol Med. 2023. PMID: 39086686 Free PMC article.
-
Loss of Cell Surface CD47 Clustering Formation and Binding Avidity to SIRPα Facilitate Apoptotic Cell Clearance by Macrophages.J Immunol. 2015 Jul 15;195(2):661-71. doi: 10.4049/jimmunol.1401719. Epub 2015 Jun 17. J Immunol. 2015. PMID: 26085683 Free PMC article.
-
Novel SIRPα Antibodies That Induce Single-Agent Phagocytosis of Tumor Cells while Preserving T Cells.J Immunol. 2021 Feb 15;206(4):712-721. doi: 10.4049/jimmunol.2001019. Epub 2021 Jan 11. J Immunol. 2021. PMID: 33431660 Free PMC article.
-
The HCMV US28 vGPCR induces potent Gαq/PLC-β signaling in monocytes leading to increased adhesion to endothelial cells.Virology. 2016 Oct;497:233-243. doi: 10.1016/j.virol.2016.07.025. Epub 2016 Aug 4. Virology. 2016. PMID: 27497185 Free PMC article.
References
-
- Ohnishi H, Kaneko Y, Okazawa H, Miyashita M, Sato R et al. (2005) Differential localization of Src homology 2 domain-containing protein tyrosine phosphatase substrate-1 and CD47 and its molecular mechanisms in cultured hippocampal neurons. J Neurosci 25: 2702-2711. doi:10.1523/JNEUROSCI.5173-04.2005. PubMed: 15758180. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
