

Mph1 and Mus81-Mms4 prevent aberrant processing of mitotic recombination intermediates
- PMID: 24119400
- PMCID: PMC3818723
- DOI: 10.1016/j.molcel.2013.09.007
Mph1 and Mus81-Mms4 prevent aberrant processing of mitotic recombination intermediates
Abstract
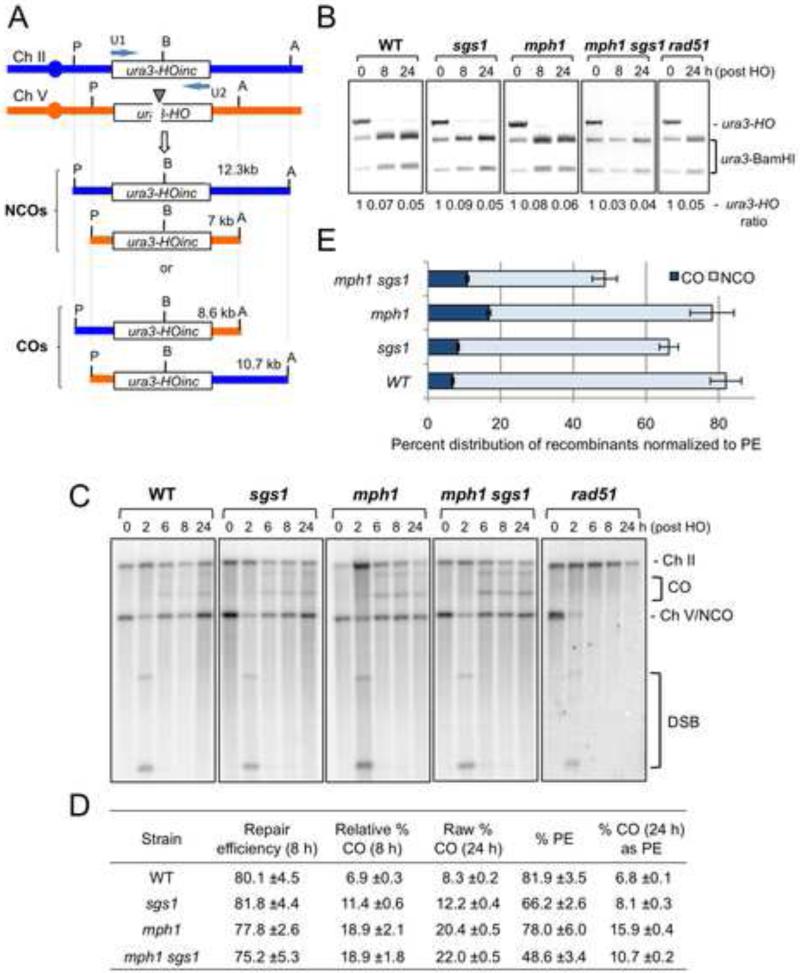
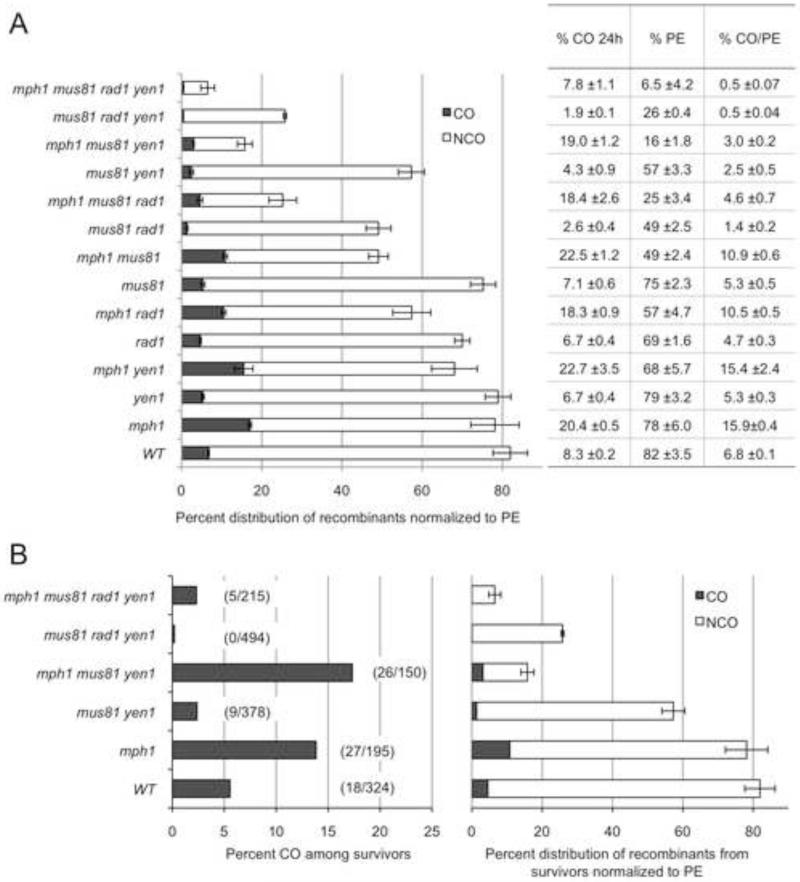
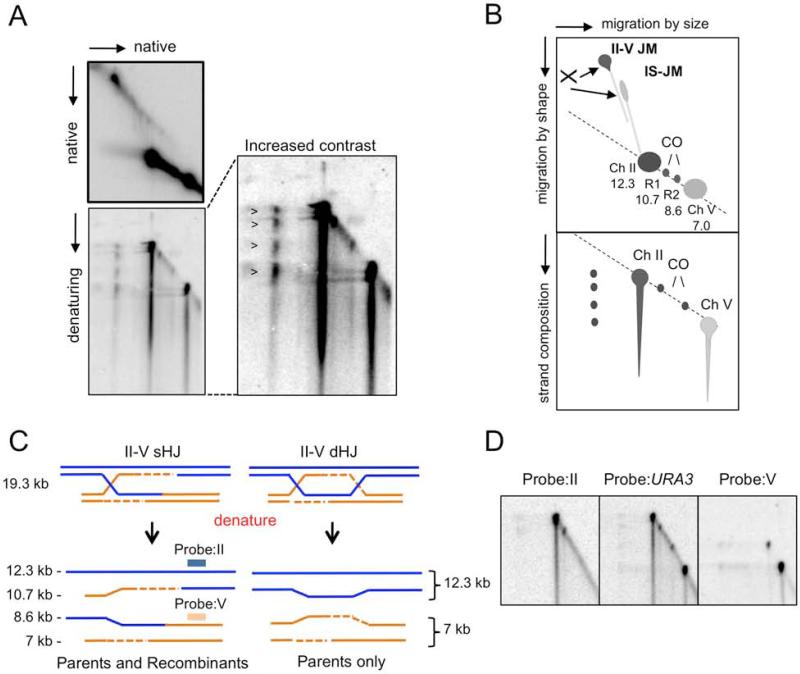
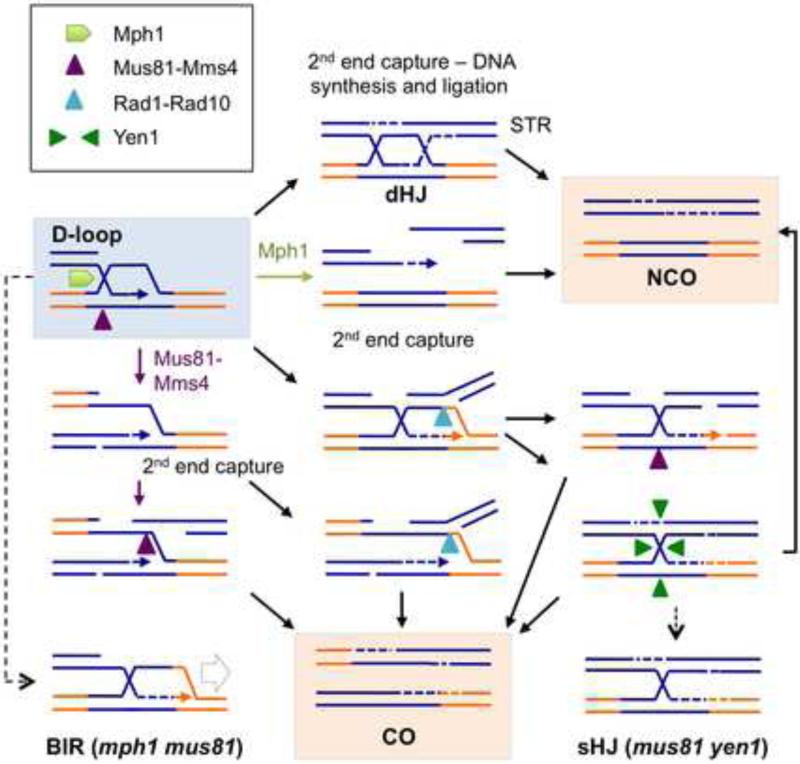
Homology-dependent repair of double-strand breaks (DSBs) from nonsister templates has the potential to generate loss of heterozygosity or genome rearrangements. Here we show that the Saccharomyces cerevisiae Mph1 helicase prevents crossovers between ectopic sequences by removing substrates for Mus81-Mms4 or Rad1-Rad10 cleavage. A role for Yen1 is only apparent in the absence of Mus81. Cells lacking Mph1 and the three nucleases are highly defective in the repair of a single DSB, suggesting that the recombination intermediates that accumulate cannot be processed by the Sgs1-Top3-Rmi1 complex (STR). Consistent with this hypothesis, ectopic joint molecules (JMs) accumulate transiently in the mph1Δ mutant and persistently when Mus81 is eliminated. Furthermore, the ectopic JMs formed in the mus81Δ mutant contain a single Holliday junction (HJ) explaining why STR is unable to process them. We suggest that Mph1 and Mus81-Mms4 recognize an early strand exchange intermediate and direct repair to noncrossover or crossover outcomes, respectively.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures


Similar articles
-
The Rad1-Rad10 nuclease promotes chromosome translocations between dispersed repeats.Nat Struct Mol Biol. 2012 Sep;19(9):964-71. doi: 10.1038/nsmb.2359. Epub 2012 Aug 12. Nat Struct Mol Biol. 2012. PMID: 22885325 Free PMC article.
-
Functional overlap between the structure-specific nucleases Yen1 and Mus81-Mms4 for DNA-damage repair in S. cerevisiae.DNA Repair (Amst). 2010 Apr 4;9(4):394-402. doi: 10.1016/j.dnarep.2009.12.017. Epub 2010 Jan 27. DNA Repair (Amst). 2010. PMID: 20106725
-
RecQ helicase, Sgs1, and XPF family endonuclease, Mus81-Mms4, resolve aberrant joint molecules during meiotic recombination.Mol Cell. 2008 Aug 8;31(3):324-36. doi: 10.1016/j.molcel.2008.07.006. Mol Cell. 2008. PMID: 18691965 Free PMC article.
-
Processing of homologous recombination repair intermediates by the Sgs1-Top3-Rmi1 and Mus81-Mms4 complexes.Cell Cycle. 2011 Sep 15;10(18):3078-85. doi: 10.4161/cc.10.18.16919. Epub 2011 Sep 15. Cell Cycle. 2011. PMID: 21876385 Review.
-
The nucleolytic resolution of recombination intermediates in yeast mitotic cells.FEMS Yeast Res. 2016 Sep;16(6):fow065. doi: 10.1093/femsyr/fow065. Epub 2016 Aug 9. FEMS Yeast Res. 2016. PMID: 27509904 Review.
Cited by
-
Pathways and Mechanisms that Prevent Genome Instability in Saccharomyces cerevisiae.Genetics. 2017 Jul;206(3):1187-1225. doi: 10.1534/genetics.112.145805. Genetics. 2017. PMID: 28684602 Free PMC article. Review.
-
Repair of a Site-Specific DNA Cleavage: Old-School Lessons for Cas9-Mediated Gene Editing.ACS Chem Biol. 2018 Feb 16;13(2):397-405. doi: 10.1021/acschembio.7b00760. Epub 2017 Nov 14. ACS Chem Biol. 2018. PMID: 29083855 Free PMC article. Review.
-
Srs2 and Mus81-Mms4 Prevent Accumulation of Toxic Inter-Homolog Recombination Intermediates.PLoS Genet. 2016 Jul 7;12(7):e1006136. doi: 10.1371/journal.pgen.1006136. eCollection 2016 Jul. PLoS Genet. 2016. PMID: 27390022 Free PMC article.
-
Homologous Recombination and the Formation of Complex Genomic Rearrangements.Trends Cell Biol. 2019 Feb;29(2):135-149. doi: 10.1016/j.tcb.2018.10.006. Epub 2018 Nov 26. Trends Cell Biol. 2019. PMID: 30497856 Free PMC article. Review.
-
Control of structure-specific endonucleases to maintain genome stability.Nat Rev Mol Cell Biol. 2017 May;18(5):315-330. doi: 10.1038/nrm.2016.177. Epub 2017 Mar 22. Nat Rev Mol Cell Biol. 2017. PMID: 28327556 Review.
References
-
- Adams MD, McVey M, Sekelsky JJ. Drosophila BLM in double-strand break repair by synthesis-dependent strand annealing. Science. 2003;299:265–267. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
