

Superior induction and maintenance of protective CD8 T cells in mice infected with mouse cytomegalovirus vector expressing RAE-1γ
- PMID: 24052528
- PMCID: PMC3799388
- DOI: 10.1073/pnas.1310215110
Superior induction and maintenance of protective CD8 T cells in mice infected with mouse cytomegalovirus vector expressing RAE-1γ
Abstract
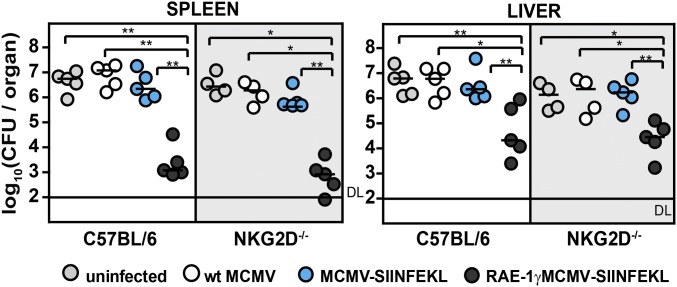
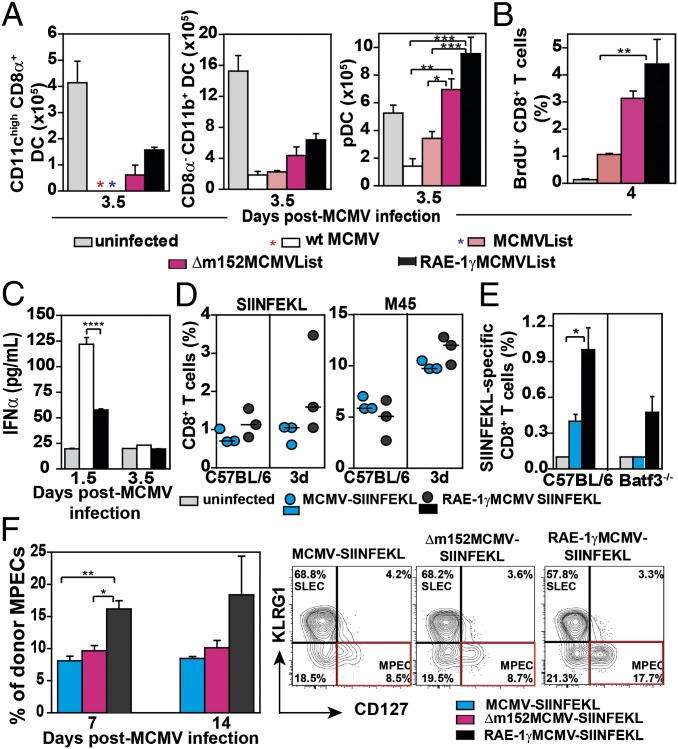
Due to a unique pattern of CD8 T-cell response induced by cytomegaloviruses (CMVs), live attenuated CMVs are attractive candidates for vaccine vectors for a number of clinically relevant infections and tumors. NKG2D is one of the most important activating NK cell receptors that plays a role in costimulation of CD8 T cells. Here we demonstrate that the expression of CD8 T-cell epitope of Listeria monocytogenes by a recombinant mouse CMV (MCMV) expressing the NKG2D ligand retinoic acid early-inducible protein 1-gamma (RAE-1γ) dramatically enhanced the effectiveness and longevity of epitope-specific CD8 T-cell response and conferred protection against a subsequent challenge infection with Listeria monocytogenes. Unexpectedly, the attenuated growth in vivo of the CMV vector expressing RAE-1γ and its capacity to enhance specific CD8 T-cell response were preserved even in mice lacking NKG2D, implying additional immune function for RAE-1γ beyond engagement of NKG2D. Thus, vectors expressing RAE-1γ represent a promising approach in the development of CD8 T-cell-based vaccines.
Keywords: CD8 T cell vaccine; RAE-1 gamma; vaccine vector.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Cytomegalovirus vector expressing RAE-1γ induces enhanced anti-tumor capacity of murine CD8+ T cells.Eur J Immunol. 2017 Aug;47(8):1354-1367. doi: 10.1002/eji.201746964. Epub 2017 Jul 4. Eur J Immunol. 2017. PMID: 28612942 Free PMC article.
-
Murine CMV Expressing the High Affinity NKG2D Ligand MULT-1: A Model for the Development of Cytomegalovirus-Based Vaccines.Front Immunol. 2018 May 7;9:991. doi: 10.3389/fimmu.2018.00991. eCollection 2018. Front Immunol. 2018. PMID: 29867968 Free PMC article.
-
Memory CD8 T Cells Generated by Cytomegalovirus Vaccine Vector Expressing NKG2D Ligand Have Effector-Like Phenotype and Distinct Functional Features.Front Immunol. 2021 Jun 3;12:681380. doi: 10.3389/fimmu.2021.681380. eCollection 2021. Front Immunol. 2021. PMID: 34168650 Free PMC article.
-
Vaccine vectors: the bright side of cytomegalovirus.Med Microbiol Immunol. 2019 Aug;208(3-4):349-363. doi: 10.1007/s00430-019-00597-7. Epub 2019 Mar 21. Med Microbiol Immunol. 2019. PMID: 30900089 Review.
-
Promising Cytomegalovirus-Based Vaccine Vector Induces Robust CD8+ T-Cell Response.Int J Mol Sci. 2019 Sep 10;20(18):4457. doi: 10.3390/ijms20184457. Int J Mol Sci. 2019. PMID: 31510028 Free PMC article. Review.
Cited by
-
Cytomegalovirus vector expressing RAE-1γ induces enhanced anti-tumor capacity of murine CD8+ T cells.Eur J Immunol. 2017 Aug;47(8):1354-1367. doi: 10.1002/eji.201746964. Epub 2017 Jul 4. Eur J Immunol. 2017. PMID: 28612942 Free PMC article.
-
Cytomegalovirus protein m154 perturbs the adaptor protein-1 compartment mediating broad-spectrum immune evasion.Elife. 2020 Jan 13;9:e50803. doi: 10.7554/eLife.50803. Elife. 2020. PMID: 31928630 Free PMC article.
-
ChAdOx1-S adenoviral vector vaccine applied intranasally elicits superior mucosal immunity compared to the intramuscular route of vaccination.Eur J Immunol. 2022 Jun;52(6):936-945. doi: 10.1002/eji.202249823. Epub 2022 Mar 31. Eur J Immunol. 2022. PMID: 35304741 Free PMC article.
-
Murine CMV Expressing the High Affinity NKG2D Ligand MULT-1: A Model for the Development of Cytomegalovirus-Based Vaccines.Front Immunol. 2018 May 7;9:991. doi: 10.3389/fimmu.2018.00991. eCollection 2018. Front Immunol. 2018. PMID: 29867968 Free PMC article.
-
NKG2D: A Master Regulator of Immune Cell Responsiveness.Front Immunol. 2018 Mar 8;9:441. doi: 10.3389/fimmu.2018.00441. eCollection 2018. Front Immunol. 2018. PMID: 29568297 Free PMC article. Review.
References
-
- Reddehase MJ. Antigens and immunoevasins: Opponents in cytomegalovirus immune surveillance. Nat Rev Immunol. 2002;2(11):831–844. - PubMed
-
- Lemmermann NA, Böhm V, Holtappels R, Reddehase MJ. In vivo impact of cytomegalovirus evasion of CD8 T-cell immunity: Facts and thoughts based on murine models. Virus Res. 2011;157(2):161–174. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
