

Analysis of the RelA:CBP/p300 interaction reveals its involvement in NF-κB-driven transcription
- PMID: 24019758
- PMCID: PMC3760798
- DOI: 10.1371/journal.pbio.1001647
Analysis of the RelA:CBP/p300 interaction reveals its involvement in NF-κB-driven transcription
Abstract
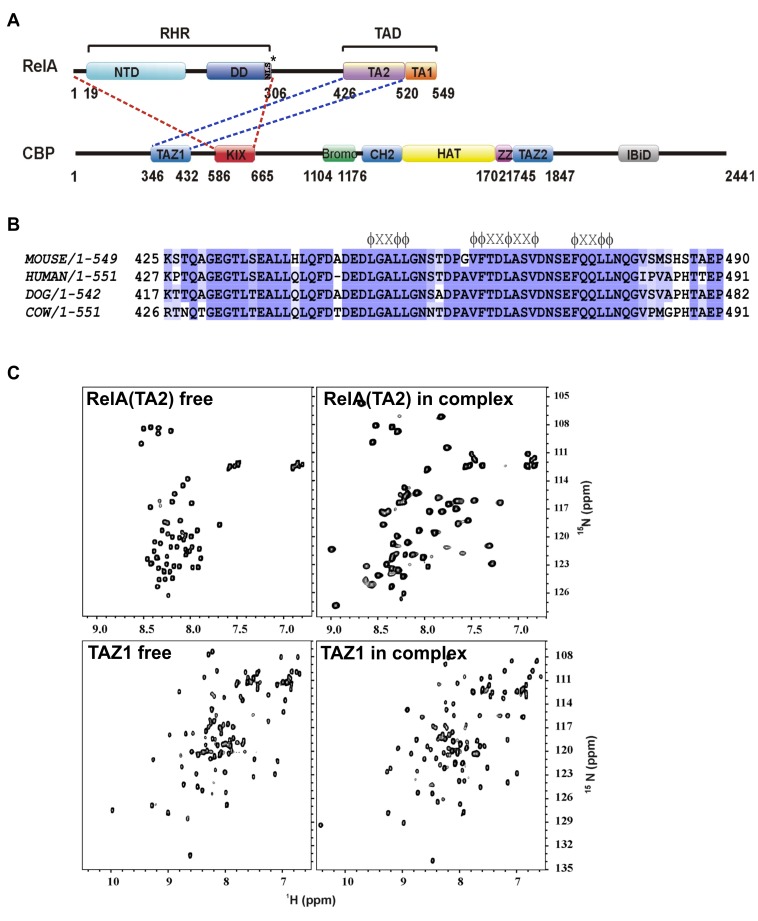
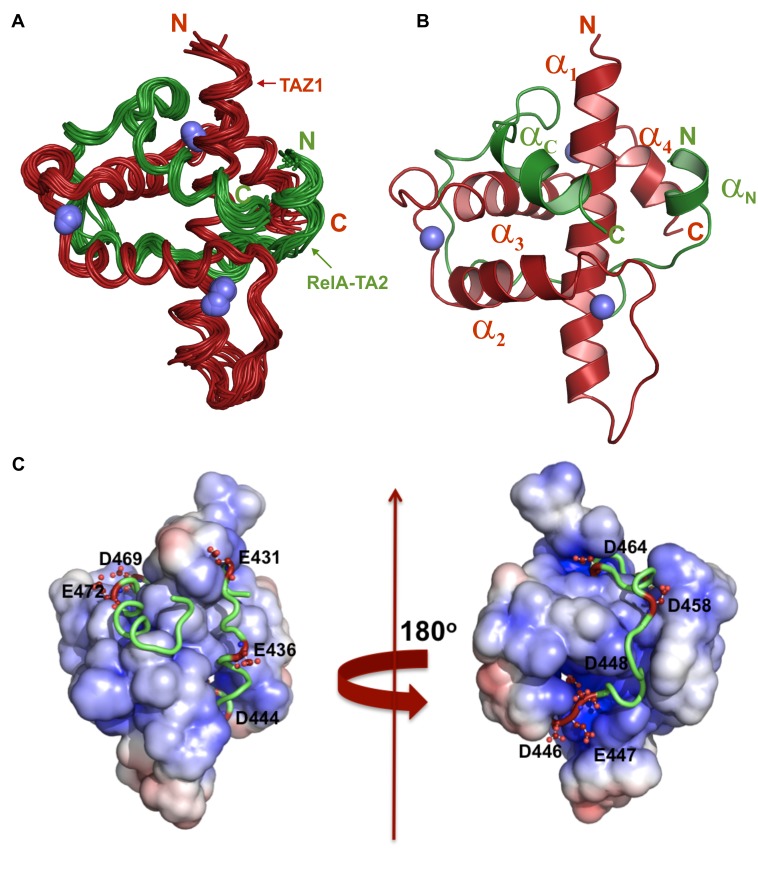
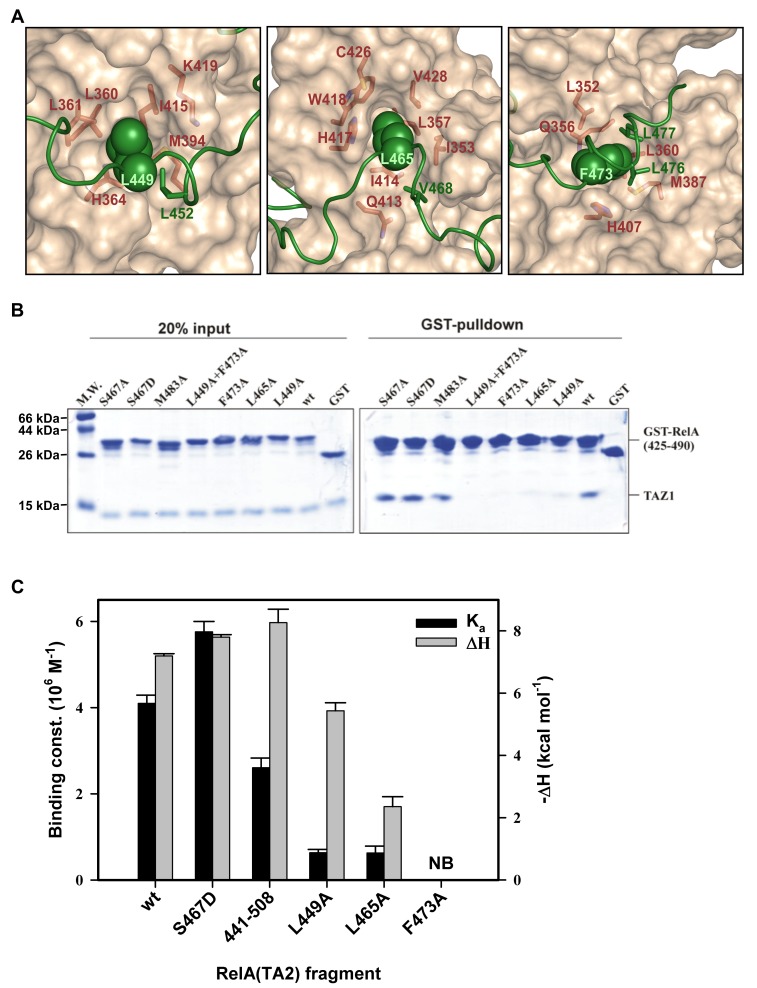
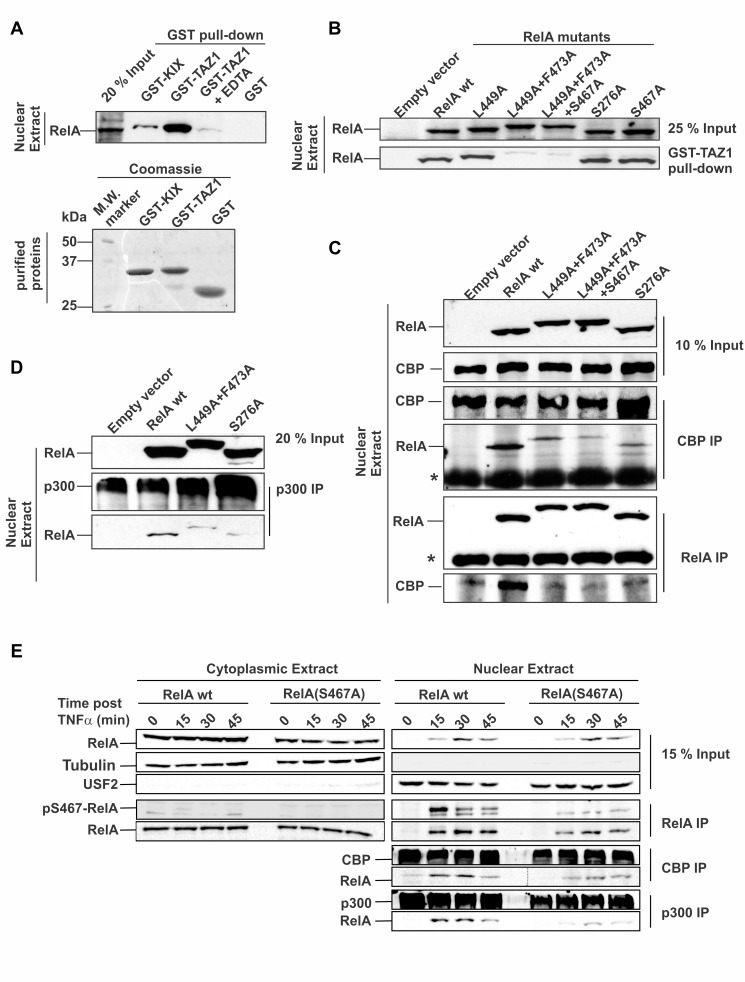
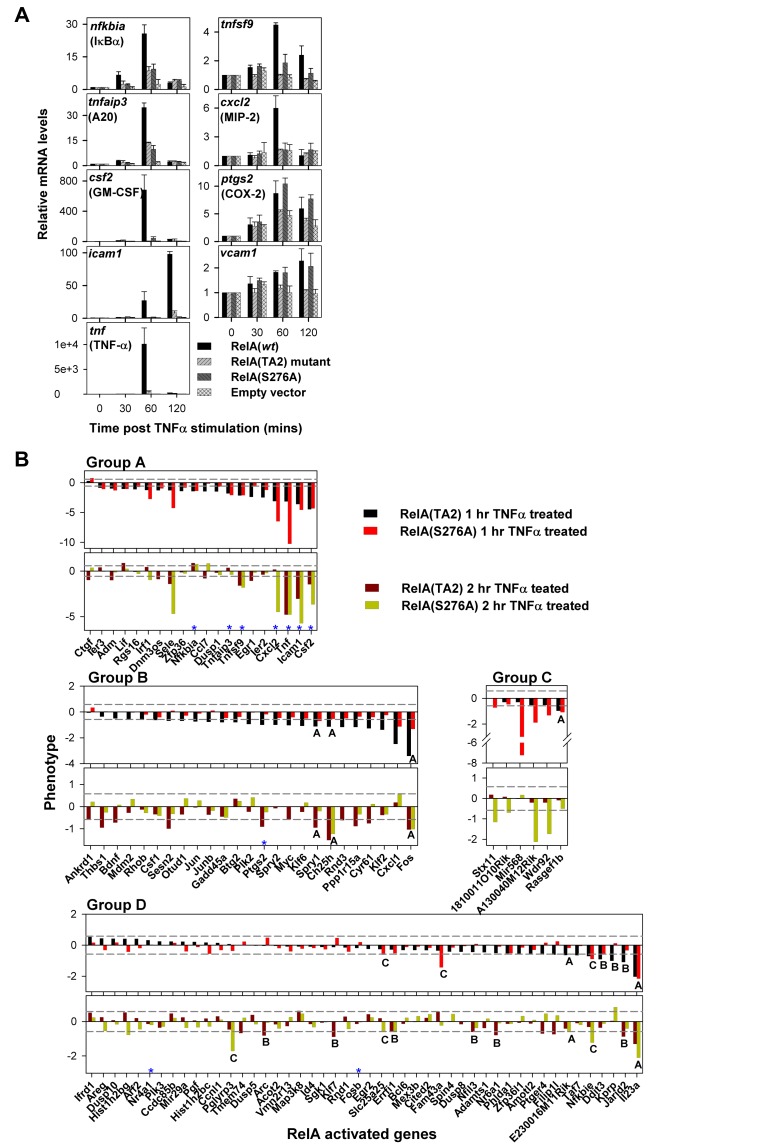
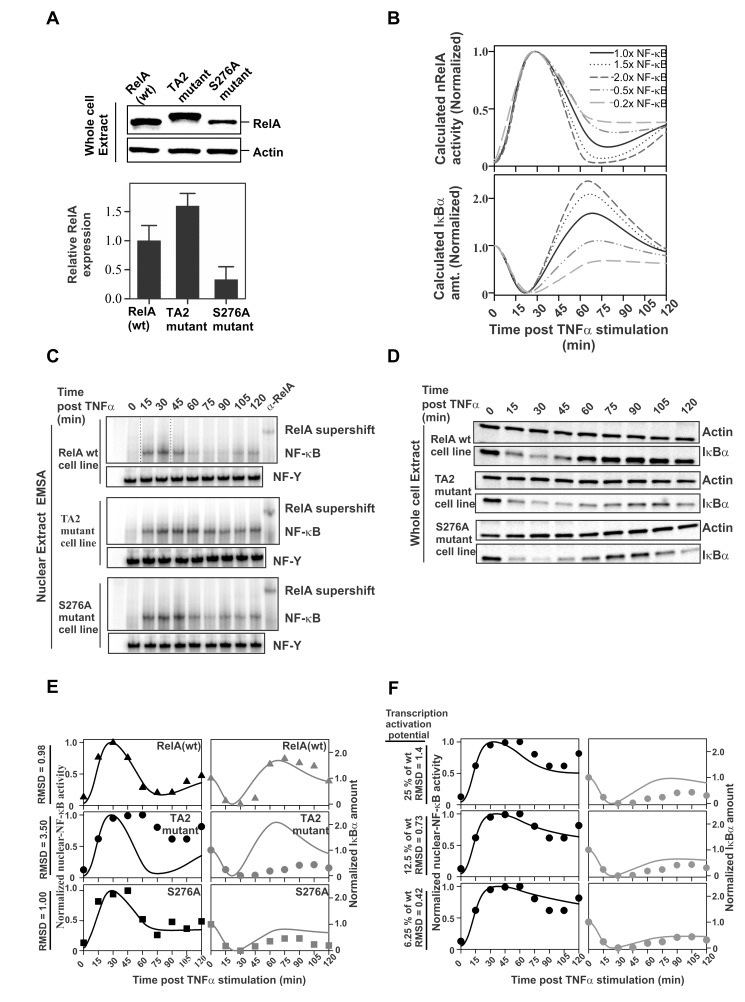
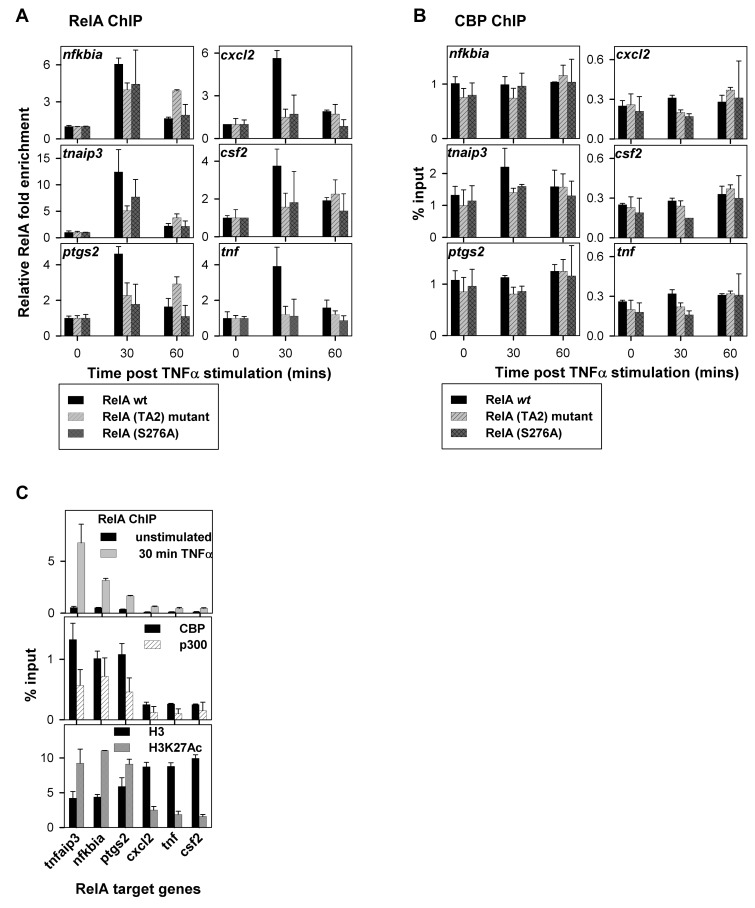
NF-κB plays a vital role in cellular immune and inflammatory response, survival, and proliferation by regulating the transcription of various genes involved in these processes. To activate transcription, RelA (a prominent NF-κB family member) interacts with transcriptional co-activators like CREB-binding protein (CBP) and its paralog p300 in addition to its cognate κB sites on the promoter/enhancer regions of DNA. The RelA:CBP/p300 complex is comprised of two components--first, DNA binding domain of RelA interacts with the KIX domain of CBP/p300, and second, the transcriptional activation domain (TAD) of RelA binds to the TAZ1 domain of CBP/p300. A phosphorylation event of a well-conserved RelA(Ser276) is prerequisite for the former interaction to occur and is considered a decisive factor for the overall RelA:CBP/p300 interaction. The role of the latter interaction in the transcription of RelA-activated genes remains unclear. Here we provide the solution structure of the latter component of the RelA:CBP complex by NMR spectroscopy. The structure reveals the folding of RelA-TA2 (a section of TAD) upon binding to TAZ1 through its well-conserved hydrophobic sites in a series of grooves on the TAZ1 surface. The structural analysis coupled with the mechanistic studies by mutational and isothermal calorimetric analyses allowed the design of RelA-mutants that selectively abrogated the two distinct components of the RelA:CBP/p300 interaction. Detailed studies of these RelA mutants using cell-based techniques, mathematical modeling, and genome-wide gene expression analysis showed that a major set of the RelA-activated genes, larger than previously believed, is affected by this interaction. We further show how the RelA:CBP/p300 interaction controls the nuclear response of NF-κB through the negative feedback loop of NF-κB pathway. Additionally, chromatin analyses of RelA target gene promoters showed constitutive recruitment of CBP/p300, thus indicating a possible role of CBP/p300 in recruitment of RelA to its target promoter sites.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Akt stimulates the transactivation potential of the RelA/p65 Subunit of NF-kappa B through utilization of the Ikappa B kinase and activation of the mitogen-activated protein kinase p38.J Biol Chem. 2001 Jun 1;276(22):18934-40. doi: 10.1074/jbc.M101103200. Epub 2001 Mar 20. J Biol Chem. 2001. PMID: 11259436
-
Differential transcriptional activation by human T-cell leukemia virus type 1 Tax mutants is mediated by distinct interactions with CREB binding protein and p300.Mol Cell Biol. 1998 Apr;18(4):2392-405. doi: 10.1128/MCB.18.4.2392. Mol Cell Biol. 1998. PMID: 9528808 Free PMC article.
-
IkappaB kinase alpha-mediated derepression of SMRT potentiates acetylation of RelA/p65 by p300.Mol Cell Biol. 2006 Jan;26(2):457-71. doi: 10.1128/MCB.26.2.457-471.2006. Mol Cell Biol. 2006. PMID: 16382138 Free PMC article.
-
Transcriptional/epigenetic regulator CBP/p300 in tumorigenesis: structural and functional versatility in target recognition.Cell Mol Life Sci. 2013 Nov;70(21):3989-4008. doi: 10.1007/s00018-012-1254-4. Epub 2013 Jan 11. Cell Mol Life Sci. 2013. PMID: 23307074 Free PMC article. Review.
-
Role of Intrinsic Protein Disorder in the Function and Interactions of the Transcriptional Coactivators CREB-binding Protein (CBP) and p300.J Biol Chem. 2016 Mar 25;291(13):6714-22. doi: 10.1074/jbc.R115.692020. Epub 2016 Feb 5. J Biol Chem. 2016. PMID: 26851278 Free PMC article. Review.
Cited by
-
The Complex of p-Tyr42 RhoA and p-p65/RelA in Response to LPS Regulates the Expression of Phosphoglycerate Kinase 1.Antioxidants (Basel). 2023 Dec 8;12(12):2090. doi: 10.3390/antiox12122090. Antioxidants (Basel). 2023. PMID: 38136210 Free PMC article.
-
LncRNA HOTAIR regulates lipopolysaccharide-induced cytokine expression and inflammatory response in macrophages.Sci Rep. 2018 Oct 23;8(1):15670. doi: 10.1038/s41598-018-33722-2. Sci Rep. 2018. PMID: 30353135 Free PMC article.
-
Transcriptional regulation in the immune system: a status report.Trends Immunol. 2014 May;35(5):190-4. doi: 10.1016/j.it.2014.03.003. Epub 2014 Apr 2. Trends Immunol. 2014. PMID: 24703179 Free PMC article.
-
DEAD-box RNA helicases with special reference to p68: Unwinding their biology, versatility, and therapeutic opportunity in cancer.Genes Dis. 2022 Mar 21;10(4):1220-1241. doi: 10.1016/j.gendis.2022.02.008. eCollection 2023 Jul. Genes Dis. 2022. PMID: 37397539 Free PMC article. Review.
-
CITED2 limits pathogenic inflammatory gene programs in myeloid cells.FASEB J. 2020 Sep;34(9):12100-12113. doi: 10.1096/fj.202000864R. Epub 2020 Jul 22. FASEB J. 2020. PMID: 32697413 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
