

The sensory neurons of touch
- PMID: 23972592
- PMCID: PMC3811145
- DOI: 10.1016/j.neuron.2013.07.051
The sensory neurons of touch
Abstract
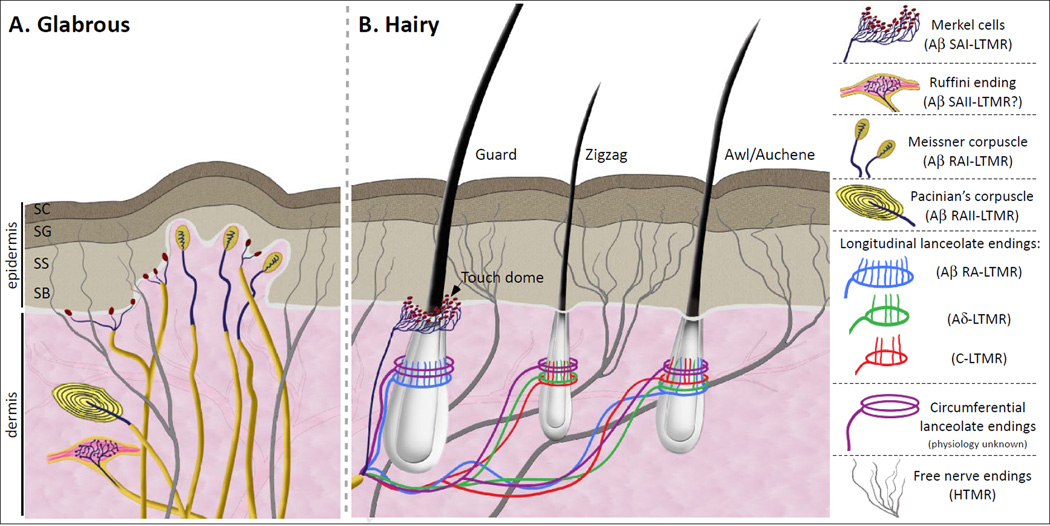
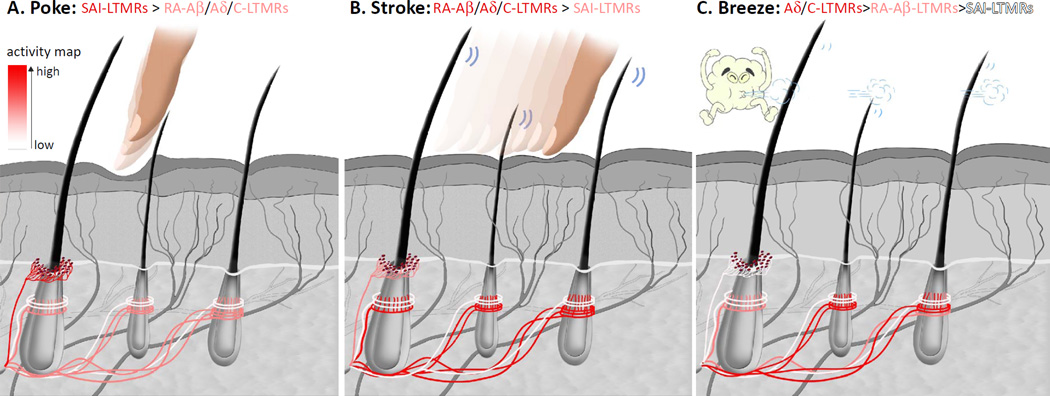
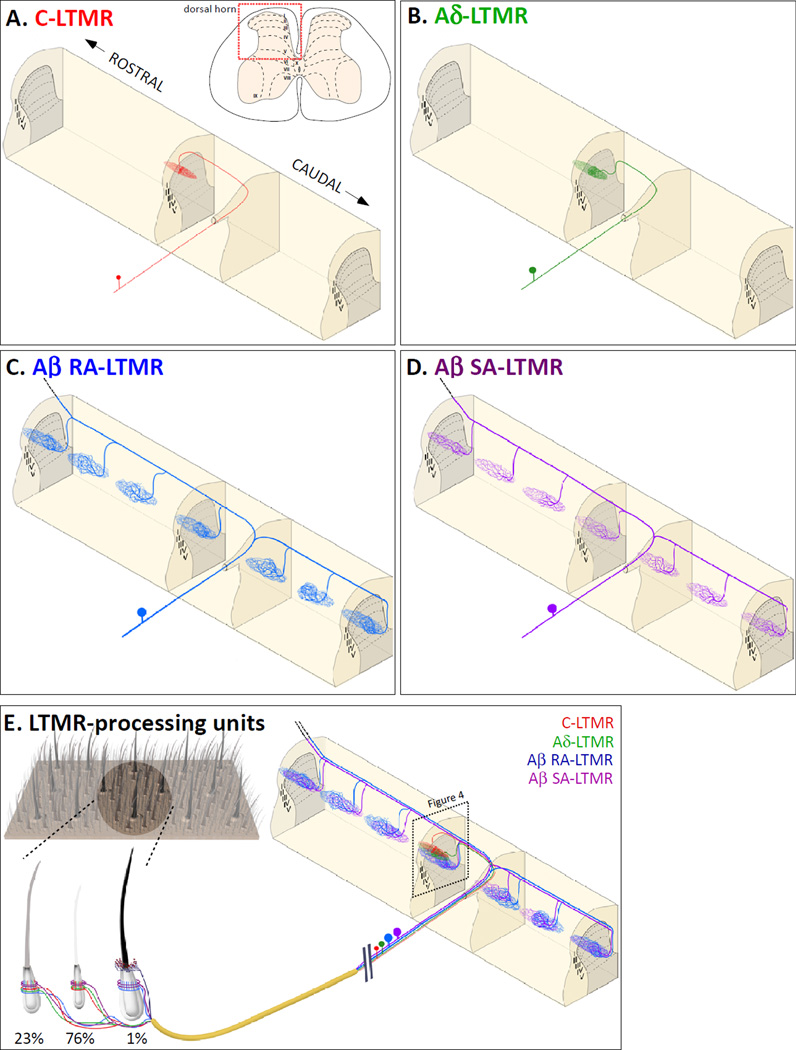
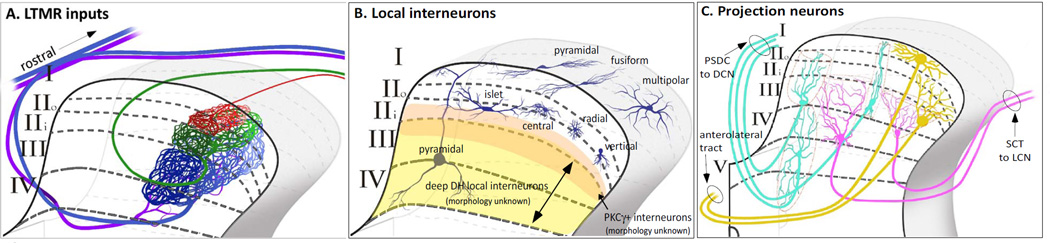
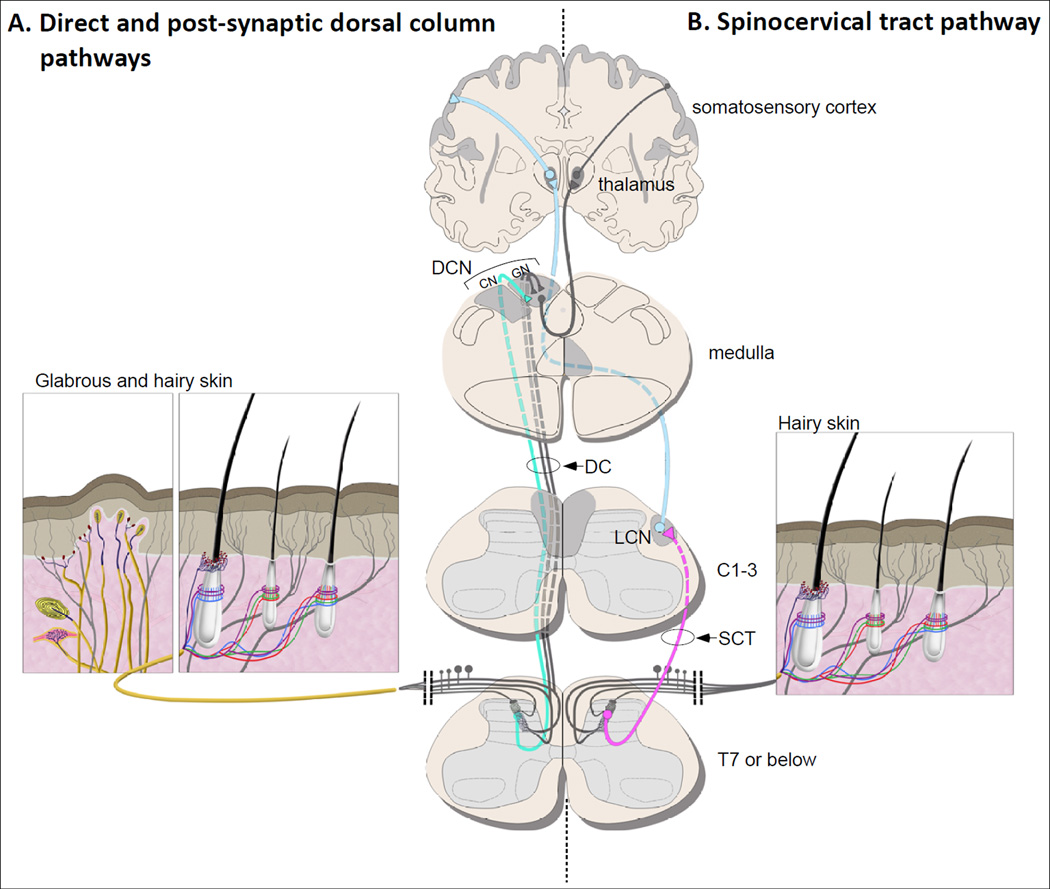
The somatosensory system decodes a wide range of tactile stimuli and thus endows us with a remarkable capacity for object recognition, texture discrimination, sensory-motor feedback and social exchange. The first step leading to perception of innocuous touch is activation of cutaneous sensory neurons called low-threshold mechanoreceptors (LTMRs). Here, we review the properties and functions of LTMRs, emphasizing the unique tuning properties of LTMR subtypes and the organizational logic of their peripheral and central axonal projections. We discuss the spinal cord neurophysiological representation of complex mechanical forces acting upon the skin and current views of how tactile information is processed and conveyed from the spinal cord to the brain. An integrative model in which ensembles of impulses arising from physiologically distinct LTMRs are integrated and processed in somatotopically aligned mechanosensory columns of the spinal cord dorsal horn underlies the nervous system's enormous capacity for perceiving the richness of the tactile world.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
The functional organization of cutaneous low-threshold mechanosensory neurons.Cell. 2011 Dec 23;147(7):1615-27. doi: 10.1016/j.cell.2011.11.027. Cell. 2011. PMID: 22196735 Free PMC article.
-
Tiling and somatotopic alignment of mammalian low-threshold mechanoreceptors.Proc Natl Acad Sci U S A. 2019 May 7;116(19):9168-9177. doi: 10.1073/pnas.1901378116. Epub 2019 Apr 17. Proc Natl Acad Sci U S A. 2019. PMID: 30996124 Free PMC article.
-
The mechanosensory neurons of touch and their mechanisms of activation.Nat Rev Neurosci. 2021 Sep;22(9):521-537. doi: 10.1038/s41583-021-00489-x. Epub 2021 Jul 26. Nat Rev Neurosci. 2021. PMID: 34312536 Free PMC article. Review.
-
Quantitative characterization of low-threshold mechanoreceptor inputs to lamina I spinoparabrachial neurons in the rat.J Physiol. 2010 Jan 1;588(Pt 1):117-24. doi: 10.1113/jphysiol.2009.181511. Epub 2009 Nov 23. J Physiol. 2010. PMID: 19933757 Free PMC article.
-
The specification and wiring of mammalian cutaneous low-threshold mechanoreceptors.Wiley Interdiscip Rev Dev Biol. 2016 May-Jun;5(3):389-404. doi: 10.1002/wdev.229. Epub 2016 Mar 17. Wiley Interdiscip Rev Dev Biol. 2016. PMID: 26992078 Free PMC article. Review.
Cited by
-
Bone marrow-derived vasculogenesis leads to scarless regeneration in deep wounds with periosteal defects.Sci Rep. 2022 Nov 29;12(1):20589. doi: 10.1038/s41598-022-24957-1. Sci Rep. 2022. PMID: 36446886 Free PMC article.
-
Synaptic circuits involving gastrin-releasing peptide receptor-expressing neurons in the dorsal horn of the mouse spinal cord.Front Mol Neurosci. 2023 Dec 7;16:1294994. doi: 10.3389/fnmol.2023.1294994. eCollection 2023. Front Mol Neurosci. 2023. PMID: 38143564 Free PMC article.
-
Thermal referral: evidence for a thermoceptive uniformity illusion without touch.Sci Rep. 2016 Oct 24;6:35286. doi: 10.1038/srep35286. Sci Rep. 2016. PMID: 27775034 Free PMC article.
-
Expression of huntingtin-associated protein 1 in adult mouse dorsal root ganglia and its neurochemical characterization in reference to sensory neuron subpopulations.IBRO Rep. 2020 Oct 6;9:258-269. doi: 10.1016/j.ibror.2020.10.001. eCollection 2020 Dec. IBRO Rep. 2020. PMID: 33089002 Free PMC article.
-
Learning touch preferences with a tactile robot using dopamine modulated STDP in a model of insular cortex.Front Neurorobot. 2015 Jul 22;9:6. doi: 10.3389/fnbot.2015.00006. eCollection 2015. Front Neurorobot. 2015. PMID: 26257639 Free PMC article.
References
-
- Adriaensen H, Gybels J, Handwerker HO, Van Hees J. Response properties of thin myelinated (A-delta) fibers in human skin nerves. Journal of neurophysiology. 1983;49:111–122. - PubMed
-
- Adrian ED. The messages in sensory nerve fibers and their interpretation. Proceedings of the Royal Society. 1931;109:1–18.
-
- Albuquerque C, Lee CJ, Jackson AC, MacDermott AB. Subpopulations of GABAergic and non-GABAergic rat dorsal horn neurons express Ca2+-permeable AMPA receptors. Eur J Neurosci. 1999;11:2758–2766. - PubMed
-
- AlChaer ED, Lawand NB, Westlund KN, Willis WD. Pelvic visceral input into the nucleus gracilis is largely mediated by the postsynaptic dorsal column pathway. Journal of neurophysiology. 1996;76:2675–2690. - PubMed
-
- Andrew D, Greenspan JD. Mechanical and heat sensitization of cutaneous nociceptors after peripheral inflammation in the rat. Journal of neurophysiology. 1999;82:2649–2656. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
