

Circularity and self-cleavage as a strategy for the emergence of a chromosome in the RNA-based protocell
- PMID: 23971788
- PMCID: PMC3765326
- DOI: 10.1186/1745-6150-8-21
Circularity and self-cleavage as a strategy for the emergence of a chromosome in the RNA-based protocell
Abstract
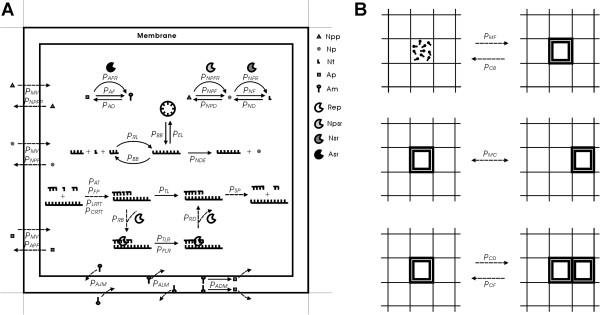
Background: It is now popularly accepted that an "RNA world" existed in early evolution. During division of RNA-based protocells, random distribution of individual genes (simultaneously as ribozymes) between offspring might have resulted in gene loss, especially when the number of gene types increased. Therefore, the emergence of a chromosome carrying linked genes was critical for the prosperity of the RNA world. However, there were quite a few immediate difficulties for this event to occur. For example, a chromosome would be much longer than individual genes, and thus more likely to degrade and less likely to replicate completely; the copying of the chromosome might start at middle sites and be only partial; and, without a complex transcription mechanism, the synthesis of distinct ribozymes would become problematic.
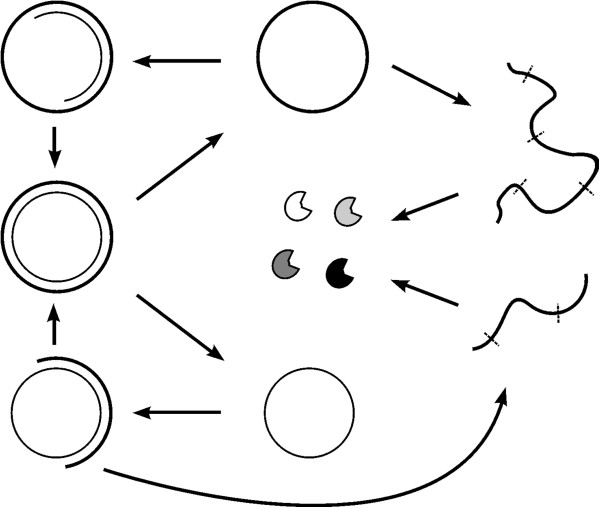
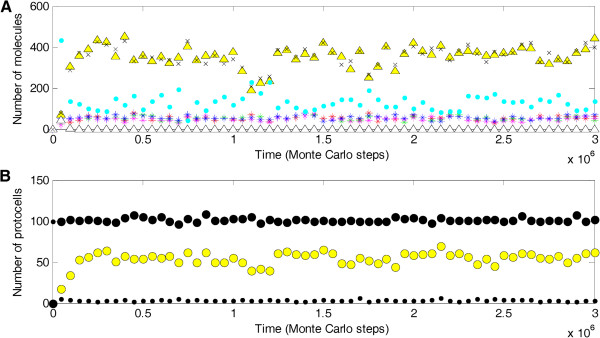
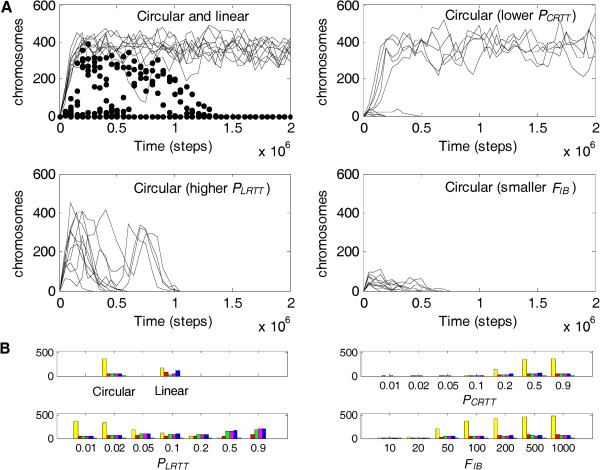
Results: Inspired by features of viroids, which have been suggested as "living fossils" of the RNA world, we supposed that these difficulties could have been overcome if the chromosome adopted a circular form and small, self-cleaving ribozymes (e.g. the hammer head ribozymes) resided at the sites between genes. Computer simulation using a Monte-Carlo method was conducted to investigate this hypothesis. The simulation shows that an RNA chromosome can spread (increase in quantity and be sustained) in the system if it is a circular one and its linear "transcripts" are readily broken at the sites between genes; the chromosome works as genetic material and ribozymes "coded" by it serve as functional molecules; and both circularity and self-cleavage are important for the spread of the chromosome.
Conclusions: In the RNA world, circularity and self-cleavage may have been adopted as a strategy to overcome the immediate difficulties for the emergence of a chromosome (with linked genes). The strategy suggested here is very simple and likely to have been used in this early stage of evolution. By demonstrating the possibility of the emergence of an RNA chromosome, this study opens on the prospect of a prosperous RNA world, populated by RNA-based protocells with a number of genes, showing complicated functions.
Figures

Similar articles
-
Evolution towards increasing complexity through functional diversification in a protocell model of the RNA world.Proc Biol Sci. 2021 Nov 24;288(1963):20212098. doi: 10.1098/rspb.2021.2098. Epub 2021 Nov 17. Proc Biol Sci. 2021. PMID: 34784760 Free PMC article.
-
Evolution of linkage and genome expansion in protocells: The origin of chromosomes.PLoS Genet. 2020 Oct 29;16(10):e1009155. doi: 10.1371/journal.pgen.1009155. eCollection 2020 Oct. PLoS Genet. 2020. PMID: 33119583 Free PMC article.
-
In search of a primitive signaling code.Biosystems. 2019 Sep;183:103984. doi: 10.1016/j.biosystems.2019.103984. Epub 2019 Jun 12. Biosystems. 2019. PMID: 31201829
-
Intramolecular RNA replicase: possibly the first self-replicating molecule in the RNA world.Orig Life Evol Biosph. 2006 Aug;36(4):413-20. doi: 10.1007/s11084-005-9006-1. Epub 2006 Aug 15. Orig Life Evol Biosph. 2006. PMID: 16909330 Review.
-
Ribozymes: Flexible molecular devices at work.Biochimie. 2011 Nov;93(11):1998-2005. doi: 10.1016/j.biochi.2011.06.026. Epub 2011 Jul 1. Biochimie. 2011. PMID: 21740954 Review.
Cited by
-
Emergence of linkage between cooperative RNA replicators encoding replication and metabolic enzymes through experimental evolution.PLoS Genet. 2023 Aug 4;19(8):e1010471. doi: 10.1371/journal.pgen.1010471. eCollection 2023 Aug. PLoS Genet. 2023. PMID: 37540715 Free PMC article.
-
Rolling Circles as a Means of Encoding Genes in the RNA World.Life (Basel). 2022 Sep 2;12(9):1373. doi: 10.3390/life12091373. Life (Basel). 2022. PMID: 36143408 Free PMC article.
-
Circular RNAs with hammerhead ribozymes encoded in eukaryotic genomes: The enemy at home.RNA Biol. 2017 Aug 3;14(8):985-991. doi: 10.1080/15476286.2017.1321730. Epub 2017 Apr 27. RNA Biol. 2017. PMID: 28448743 Free PMC article.
-
The emergence of DNA in the RNA world: an in silico simulation study of genetic takeover.BMC Evol Biol. 2015 Dec 7;15:272. doi: 10.1186/s12862-015-0548-1. BMC Evol Biol. 2015. PMID: 26643199 Free PMC article.
-
From molecular to cellular form: modeling the first major transition during the arising of life.BMC Evol Biol. 2019 Apr 3;19(1):84. doi: 10.1186/s12862-019-1412-5. BMC Evol Biol. 2019. PMID: 30943915 Free PMC article.
References
-
- Gilbert W. The RNA world. Nature. 1986;319:618.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
