

Effect of charged residues in the N-domain of Sup35 protein on prion [PSI+] stability and propagation
- PMID: 23965990
- PMCID: PMC3789951
- DOI: 10.1074/jbc.M113.471805
Effect of charged residues in the N-domain of Sup35 protein on prion [PSI+] stability and propagation
Abstract
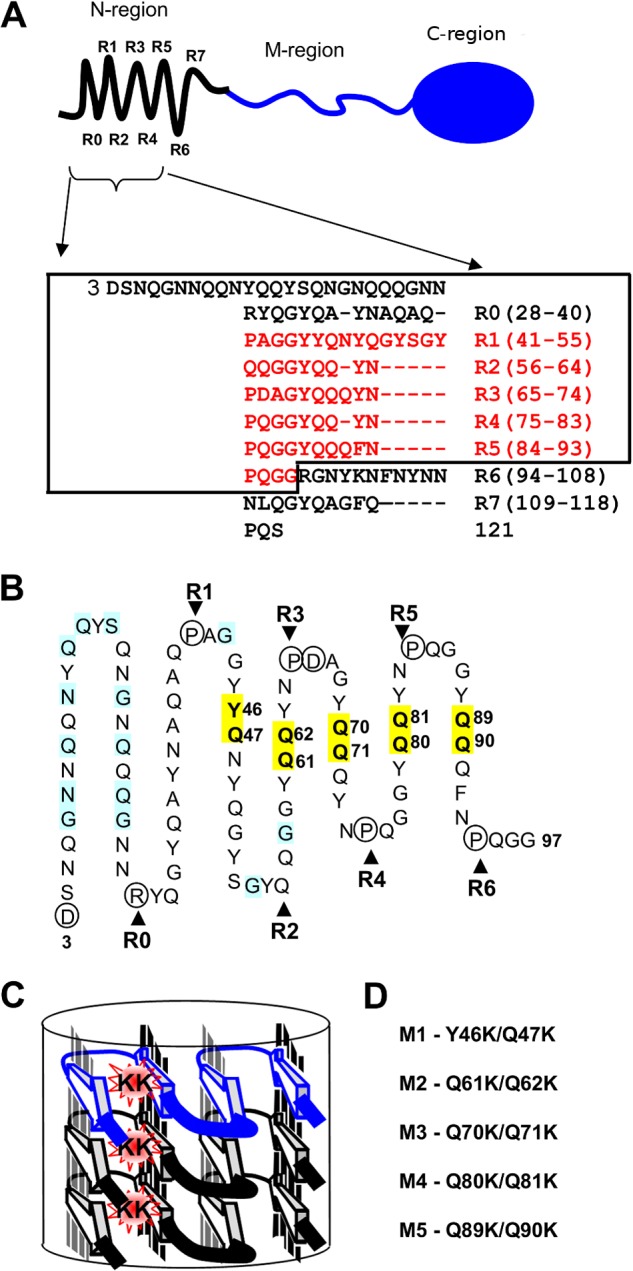
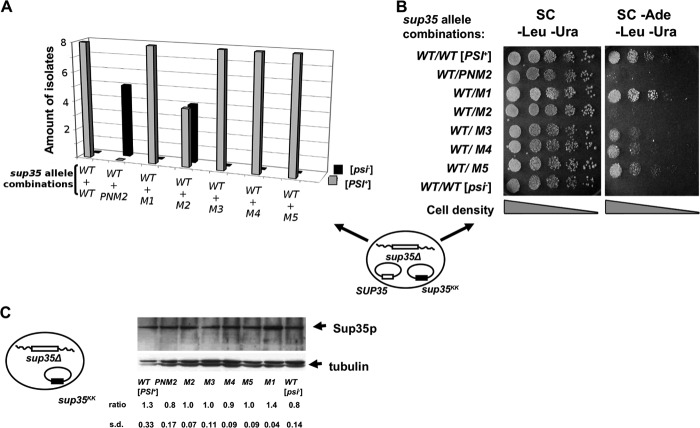
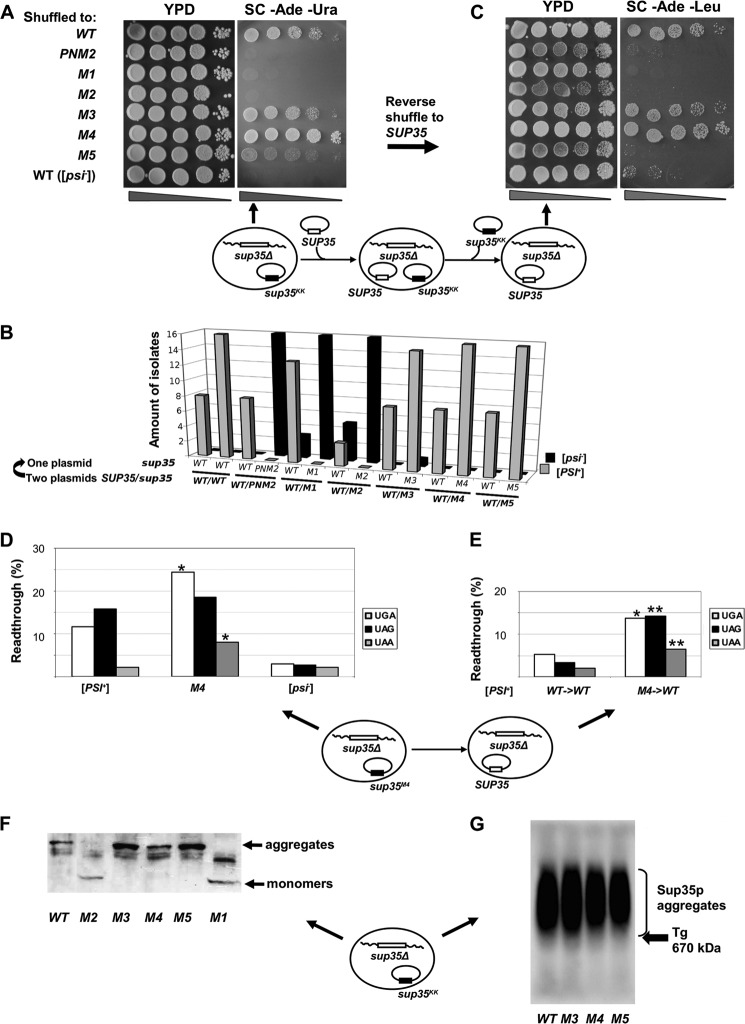
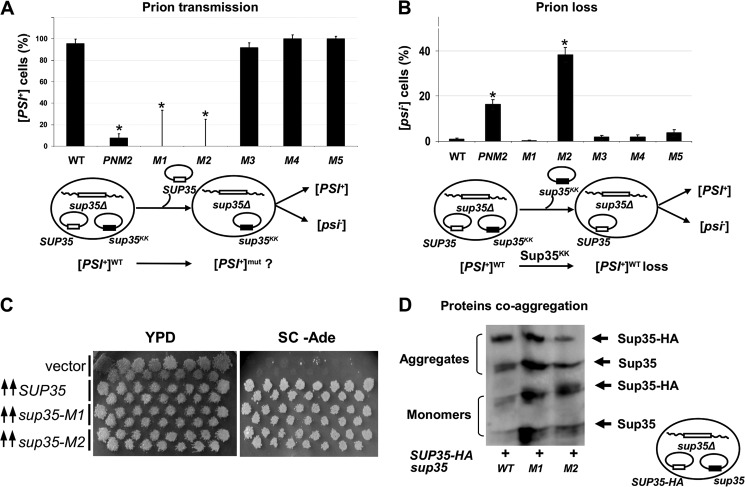
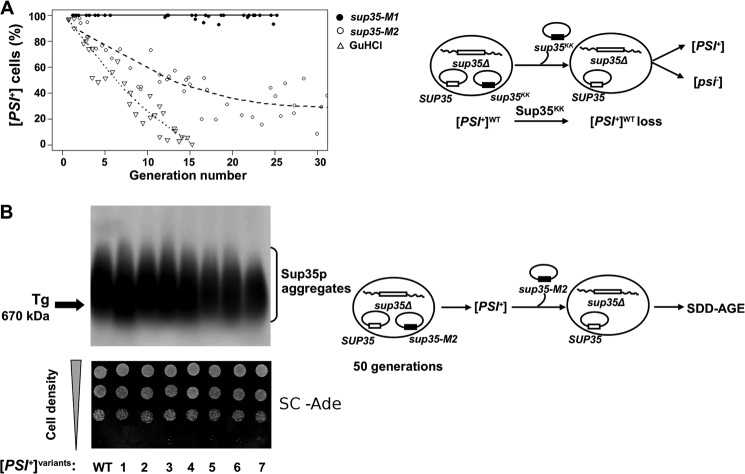
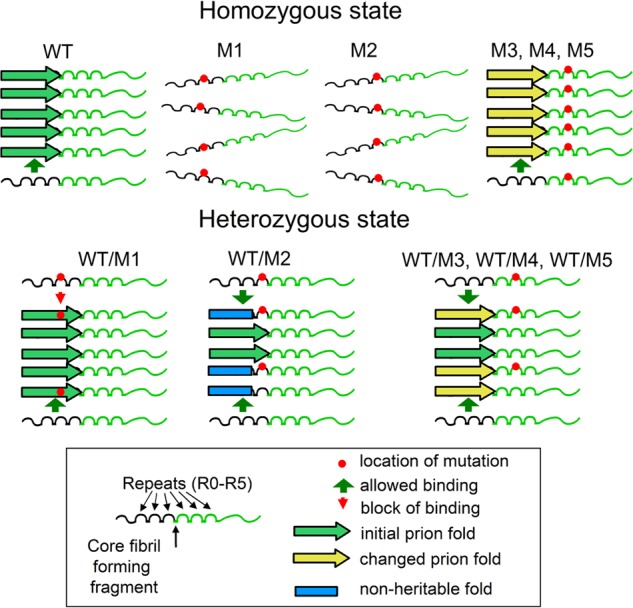
Recent studies have shown that Sup35p prion fibrils probably have a parallel in-register β-structure. However, the part(s) of the N-domain critical for fibril formation and maintenance of the [PSI(+)] phenotype remains unclear. Here we designed a set of five SUP35 mutant alleles (sup35(KK)) with lysine substitutions in each of five N-domain repeats, and investigated their effect on infectivity and ability of corresponding proteins to aggregate and coaggregate with wild type Sup35p in the [PSI(+)] strain. Alleles sup35-M1 (Y46K/Q47K) and sup35-M2 (Q61K/Q62K) led to prion loss, whereas sup35-M3 (Q70K/Q71K), sup35-M4 (Q80K/Q81K), and sup35-M5 (Q89K/Q90K) were able to maintain the [PSI(+)] prion. This suggests that the critical part of the parallel in-register β-structure for the studied [PSI(+)] prion variant lies in the first 63-69 residues. Our study also reveals an unexpected interplay between the wild type Sup35p and proteins expressed from the sup35(KK) alleles during prionization. Both Sup35-M1p and Sup35-M2p coaggregated with Sup35p, but only sup35-M2 led to prion loss in a dominant manner. We suggest that in the fibrils, Sup35p can bind to Sup35-M1p in the same conformation, whereas Sup35-M2p only allowed the Sup35p conformation that leads to the non-heritable fold. Mutations sup35-M4 and sup35-M5 influence the structure of the prion forming region to a lesser extent, and can lead to the formation of new prion variants.
Keywords: Amyloid; Prions; Protein Misfolding; Protein Structure; Saccharomyces cerevisiae; Superpleated β-Structure; Translation Release Factors; Translation Termination; [PSI+].
Figures
Similar articles
-
Structure-based view on [PSI(+)] prion properties.Prion. 2015;9(3):190-9. doi: 10.1080/19336896.2015.1044186. Prion. 2015. PMID: 26030475 Free PMC article. Review.
-
[Modification of [PSI+] prion properties by the combination of amino acid changes within Sup35 protein N-domain].Mol Biol (Mosk). 2014 Mar-Apr;48(2):314-21. Mol Biol (Mosk). 2014. PMID: 25850301 Russian.
-
[PSI+] maintenance is dependent on the composition, not primary sequence, of the oligopeptide repeat domain.PLoS One. 2011;6(7):e21953. doi: 10.1371/journal.pone.0021953. Epub 2011 Jul 8. PLoS One. 2011. PMID: 21760933 Free PMC article.
-
[Mechanism and application of molecular self-assembly in Sup35 prion domain of Saccharomyces cerevisiae].Sheng Wu Gong Cheng Xue Bao. 2011 Oct;27(10):1401-7. Sheng Wu Gong Cheng Xue Bao. 2011. PMID: 22260056 Review. Chinese.
-
[Rnq1 protein protects [PSI^(+)] prion from effect of the PNM mutation].Mol Biol (Mosk). 2017 Mar-Apr;51(2):367-371. doi: 10.7868/S0026898417010050. Mol Biol (Mosk). 2017. PMID: 28537243 Russian.
Cited by
-
Distinct amino acid compositional requirements for formation and maintenance of the [PSI⁺] prion in yeast.Mol Cell Biol. 2015 Mar;35(5):899-911. doi: 10.1128/MCB.01020-14. Epub 2014 Dec 29. Mol Cell Biol. 2015. PMID: 25547291 Free PMC article.
-
How Big Is the Yeast Prion Universe?Int J Mol Sci. 2023 Jul 19;24(14):11651. doi: 10.3390/ijms241411651. Int J Mol Sci. 2023. PMID: 37511408 Free PMC article. Review.
-
Amyloid fibril length distribution from dynamic light scattering data.Eur Biophys J. 2022 Jul;51(4-5):325-333. doi: 10.1007/s00249-022-01600-5. Epub 2022 May 11. Eur Biophys J. 2022. PMID: 35546203
-
Design of a New [PSI +]-No-More Mutation in SUP35 With Strong Inhibitory Effect on the [PSI +] Prion Propagation.Front Mol Neurosci. 2019 Nov 19;12:274. doi: 10.3389/fnmol.2019.00274. eCollection 2019. Front Mol Neurosci. 2019. PMID: 31803017 Free PMC article.
-
Structure-based view on [PSI(+)] prion properties.Prion. 2015;9(3):190-9. doi: 10.1080/19336896.2015.1044186. Prion. 2015. PMID: 26030475 Free PMC article. Review.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
