

Functional diversification of Argonautes in nematodes: an expanding universe
- PMID: 23863149
- PMCID: PMC3782831
- DOI: 10.1042/BST20130086
Functional diversification of Argonautes in nematodes: an expanding universe
Abstract
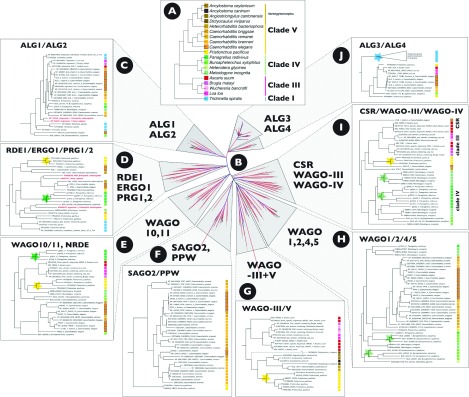
In the last decade, many diverse RNAi (RNA interference) pathways have been discovered that mediate gene silencing at epigenetic, transcriptional and post-transcriptional levels. The diversity of RNAi pathways is inherently linked to the evolution of Ago (Argonaute) proteins, the central protein component of RISCs (RNA-induced silencing complexes). An increasing number of diverse Agos have been identified in different species. The functions of most of these proteins are not yet known, but they are generally assumed to play roles in development, genome stability and/or protection against viruses. Recent research in the nematode Caenorhabditis elegans has expanded the breadth of RNAi functions to include transgenerational epigenetic memory and, possibly, environmental sensing. These functions are inherently linked to the production of secondary siRNAs (small interfering RNAs) that bind to members of a clade of WAGOs (worm-specific Agos). In the present article, we review briefly what is known about the evolution and function of Ago proteins in eukaryotes, including the expansion of WAGOs in nematodes. We postulate that the rapid evolution of WAGOs enables the exceptional functional plasticity of nematodes, including their capacity for parasitism.
Figures
Similar articles
-
Analysis of the C. elegans Argonaute family reveals that distinct Argonautes act sequentially during RNAi.Cell. 2006 Nov 17;127(4):747-57. doi: 10.1016/j.cell.2006.09.033. Cell. 2006. PMID: 17110334
-
The evolution of animal Argonautes: evidence for the absence of antiviral AGO Argonautes in vertebrates.Sci Rep. 2017 Aug 23;7(1):9230. doi: 10.1038/s41598-017-08043-5. Sci Rep. 2017. PMID: 28835645 Free PMC article.
-
The Caenorhabditis elegans HEN1 ortholog, HENN-1, methylates and stabilizes select subclasses of germline small RNAs.PLoS Genet. 2012;8(4):e1002617. doi: 10.1371/journal.pgen.1002617. Epub 2012 Apr 19. PLoS Genet. 2012. PMID: 22548001 Free PMC article.
-
Small RNAs in parasitic nematodes - forms and functions.Parasitology. 2020 Jul;147(8):855-864. doi: 10.1017/S0031182019001689. Epub 2019 Dec 17. Parasitology. 2020. PMID: 31843030 Free PMC article. Review.
-
RNAa in action: from the exception to the norm.RNA Biol. 2014;11(10):1221-5. doi: 10.4161/15476286.2014.972853. RNA Biol. 2014. PMID: 25602906 Free PMC article. Review.
Cited by
-
Key Mechanistic Principles and Considerations Concerning RNA Interference.Front Plant Sci. 2020 Aug 13;11:1237. doi: 10.3389/fpls.2020.01237. eCollection 2020. Front Plant Sci. 2020. PMID: 32903622 Free PMC article. Review.
-
Nematode endogenous small RNA pathways.Worm. 2014 Mar 5;3:e28234. doi: 10.4161/worm.28234. eCollection 2014. Worm. 2014. PMID: 25340013 Free PMC article. Review.
-
Metagenomic sequencing suggests a diversity of RNA interference-like responses to viruses across multicellular eukaryotes.PLoS Genet. 2018 Jul 30;14(7):e1007533. doi: 10.1371/journal.pgen.1007533. eCollection 2018 Jul. PLoS Genet. 2018. PMID: 30059538 Free PMC article.
-
Comparative functional characterization of the CSR-1 22G-RNA pathway in Caenorhabditis nematodes.Nucleic Acids Res. 2015 Jan;43(1):208-24. doi: 10.1093/nar/gku1308. Epub 2014 Dec 15. Nucleic Acids Res. 2015. PMID: 25510497 Free PMC article.
-
The genome of the yellow potato cyst nematode, Globodera rostochiensis, reveals insights into the basis of parasitism and virulence.Genome Biol. 2016 Jun 10;17(1):124. doi: 10.1186/s13059-016-0985-1. Genome Biol. 2016. PMID: 27286965 Free PMC article.
References
-
- Fire A., Xu S., Montgomery M.K., Kostas S.A., Driver S.E., Mello C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 1998;391:806–811. - PubMed
-
- Metzlaff M., O’Dell M., Cluster P.D., Flavell R.B. RNA-mediated RNA degradation and chalcone synthase A silencing in petunia. Cell. 1997;88:845–854. - PubMed
-
- Chapman E.J., Carrington J.C. Specialization and evolution of endogenous small RNA pathways. Nat. Rev. Genet. 2007;8:884–896. - PubMed
-
- Peters L., Meister G. Argonaute proteins: mediators of RNA silencing. Mol. Cell. 2007;26:611–623. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources