

Using EAE to better understand principles of immune function and autoimmune pathology
- PMID: 23849779
- PMCID: PMC3963137
- DOI: 10.1016/j.jaut.2013.06.008
Using EAE to better understand principles of immune function and autoimmune pathology
Abstract
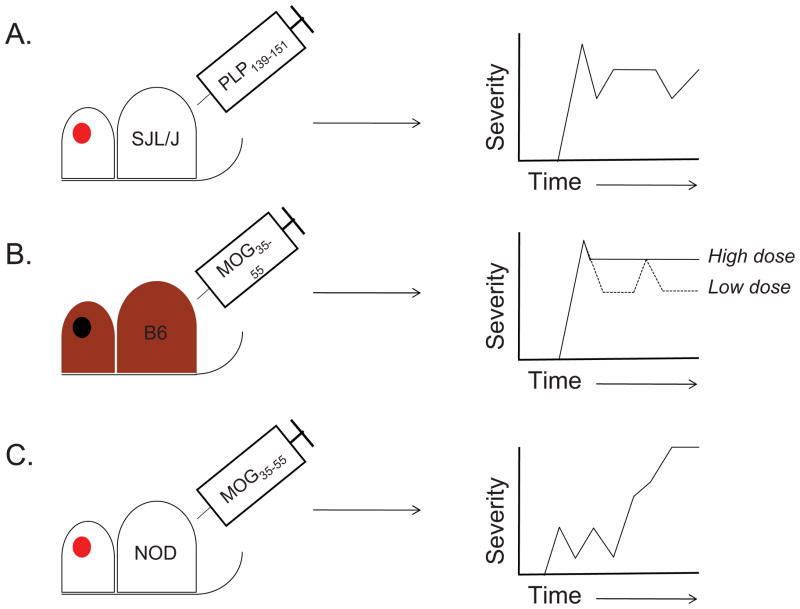
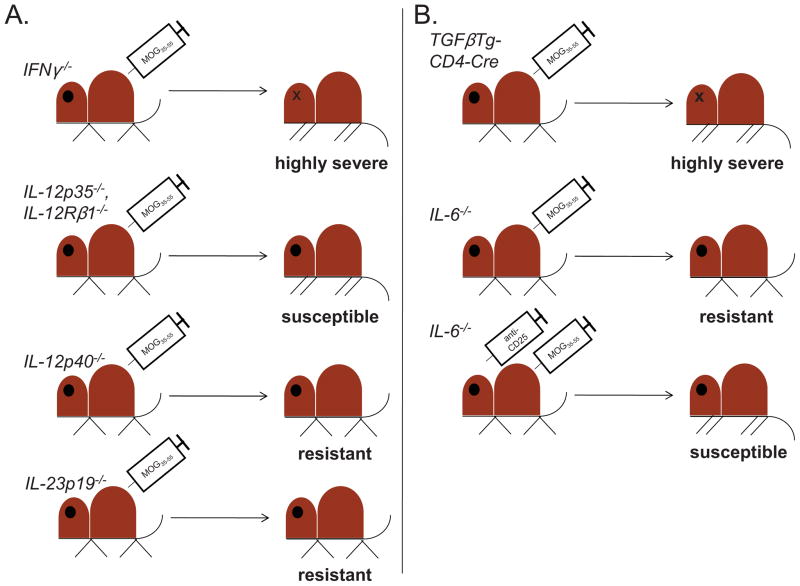
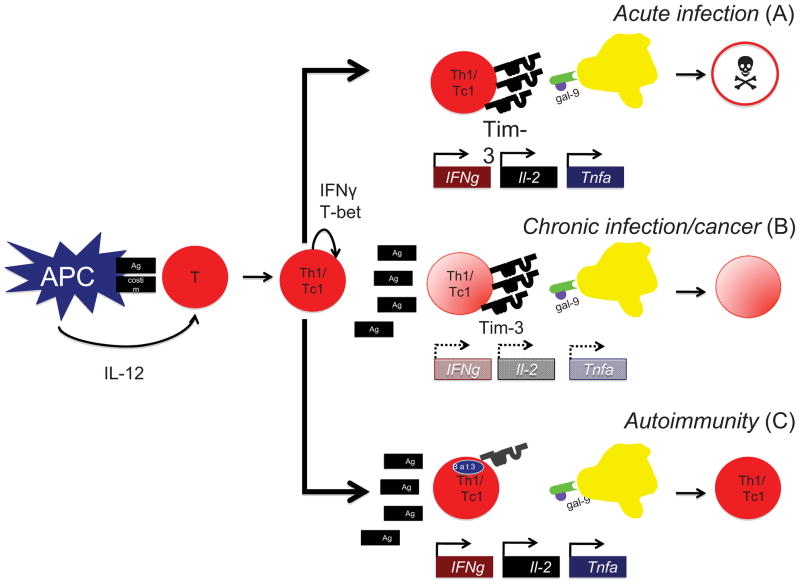
Multiple sclerosis (MS) is a chronic inflammatory disease of the central nervous system (CNS) in which myelin becomes the target of attack by autoreactive T cells. The immune components of the disease are recapitulated in mice using the experimental autoimmune encephalomyelitis (EAE) model. EAE is classically induced by the immunization of mice with encephalitogenic antigens derived from CNS proteins such as proteolipid protein (PLP), myelin basic protein (MBP) and myelin oligodendrocyte glycoprotein (MOG). Immunization of susceptible mouse strains with these antigens will induce autoreactive inflammatory T cell infiltration of the CNS. More recently, the advent of clonal T cell receptor transgenic mice has led to the development of adoptive transfer protocols in which myelin-specific T cells may induce disease upon transfer into naïve recipient animals. When used in concert with gene knockout strains, these protocols are powerful tools by which to dissect the molecular pathways that promote inflammatory T cells responses in the central nervous system (CNS). Further, myelin-antigen-specific transgenic T cells may be cultured in vitro under a variety of conditions prior to adoptive transfer, allowing one to study the effects of soluble factors or pharmacologic compounds on T cell pathogenicity. In this review, we describe many of the existing models of EAE, and discuss the contributions that use of these models has made in understanding both T helper cell differentiation and the function of inhibitory T cell receptors. We focus on the step-by-step elucidation of the network of signals required for T helper 17 (Th17) cell differentiation, as well as the molecular dissection of the Tim-3 negative regulatory signaling pathway in Th1 cells.
Keywords: Bat3; CD4+ T lymphocyte, myelin; Experimental autoimmune encephalomyelitis; Interleukin-17; Multiple sclerosis; Tim-3.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Role of Th17 cells in the pathogenesis of CNS inflammatory demyelination.J Neurol Sci. 2013 Oct 15;333(1-2):76-87. doi: 10.1016/j.jns.2013.03.002. Epub 2013 Apr 8. J Neurol Sci. 2013. PMID: 23578791 Free PMC article. Review.
-
LFA-1 Controls Th1 and Th17 Motility Behavior in the Inflamed Central Nervous System.Front Immunol. 2019 Oct 18;10:2436. doi: 10.3389/fimmu.2019.02436. eCollection 2019. Front Immunol. 2019. PMID: 31681316 Free PMC article.
-
The influence of T cell Ig mucin-3 signaling on central nervous system autoimmune disease is determined by the effector function of the pathogenic T cells.J Immunol. 2013 May 15;190(10):4991-9. doi: 10.4049/jimmunol.1300083. Epub 2013 Apr 5. J Immunol. 2013. PMID: 23562810 Free PMC article.
-
MOG extracellular domain (p1-125) triggers elevated frequency of CXCR3+ CD4+ Th1 cells in the CNS of mice and induces greater incidence of severe EAE.Mult Scler. 2014 Sep;20(10):1312-21. doi: 10.1177/1352458514524086. Epub 2014 Feb 19. Mult Scler. 2014. PMID: 24552747
-
Autoimmune pathogenesis of multiple sclerosis: role of autoreactive T lymphocytes and new immunotherapeutic strategies.Crit Rev Immunol. 1997;17(1):33-75. doi: 10.1615/critrevimmunol.v17.i1.20. Crit Rev Immunol. 1997. PMID: 9034723 Review.
Cited by
-
Platelets in Multiple Sclerosis: Early and Central Mediators of Inflammation and Neurodegeneration and Attractive Targets for Molecular Imaging and Site-Directed Therapy.Front Immunol. 2021 Feb 19;12:620963. doi: 10.3389/fimmu.2021.620963. eCollection 2021. Front Immunol. 2021. PMID: 33679764 Free PMC article. Review.
-
Tuning the Reactivity of Nuclear Factor Erythroid 2-Related Factor 2 (Nrf2) Activators for Optimal in Vivo Efficacy.ACS Med Chem Lett. 2023 Nov 21;14(12):1700-1706. doi: 10.1021/acsmedchemlett.3c00336. eCollection 2023 Dec 14. ACS Med Chem Lett. 2023. PMID: 38116420 Free PMC article.
-
IFN-β Facilitates Neuroantigen-Dependent Induction of CD25+ FOXP3+ Regulatory T Cells That Suppress Experimental Autoimmune Encephalomyelitis.J Immunol. 2016 Oct 15;197(8):2992-3007. doi: 10.4049/jimmunol.1500411. Epub 2016 Sep 12. J Immunol. 2016. PMID: 27619998 Free PMC article.
-
Regulatory B and T lymphocytes in multiple sclerosis: friends or foes?Auto Immun Highlights. 2018 Nov 10;9(1):9. doi: 10.1007/s13317-018-0109-x. Auto Immun Highlights. 2018. PMID: 30415321 Free PMC article. Review.
-
Mucosal-Associated Invariant T Cells in Multiple Sclerosis: The Jury is Still Out.Front Immunol. 2015 Sep 30;6:503. doi: 10.3389/fimmu.2015.00503. eCollection 2015. Front Immunol. 2015. PMID: 26483793 Free PMC article. Review.
References
-
- Frohman EM, Racke MK, Raine CS. Multiple sclerosis--the plaque and its pathogenesis. N Engl J Med. 2006;354:942–55. - PubMed
-
- Meuth SG, Gobel K, Wiendl H. Immune therapy of multiple sclerosis--future strategies. Curr Pharm Des. 2012;18:4489–97. - PubMed
-
- Richards TL, Alvord EC, Jr, He Y, Petersen K, Peterson J, Cosgrove S, Heide AC, Marro K, Rose LM. Experimental allergic encephalomyelitis in non-human primates: diffusion imaging of acute and chronic brain lesions. Mult Scler. 1995;1:109–17. - PubMed
-
- Aritake K, Koh CS, Inoue A, Yabuuchi F, Kitagaki K, Ikoma Y, Hayashi S. Effects of human recombinant-interferon beta in experimental autoimmune encephalomyelitis in guinea pigs. Pharm Biol. 2010;48:1273–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
